

Review
doi: 10.1038/nrm3902.
Epub 2014 Nov 5.
Extracellular matrix assembly: a multiscale deconstruction
Affiliations
- PMID: 25370693
- PMCID: PMC4682873
- DOI: 10.1038/nrm3902
Item in Clipboard
Review
Extracellular matrix assembly: a multiscale deconstruction
Nat Rev Mol Cell Biol.
2014 Dec.
Abstract
The biochemical and biophysical properties of the extracellular matrix (ECM) dictate tissue-specific cell behaviour. The molecules that are associated with the ECM of each tissue, including collagens, proteoglycans, laminins and fibronectin, and the manner in which they are assembled determine the structure and the organization of the resultant ECM. The product is a specific ECM signature that is comprised of unique compositional and topographical features that both reflect and facilitate the functional requirements of the tissue.
Conflict of interest statement
The authors declare no competing interests.
Figures
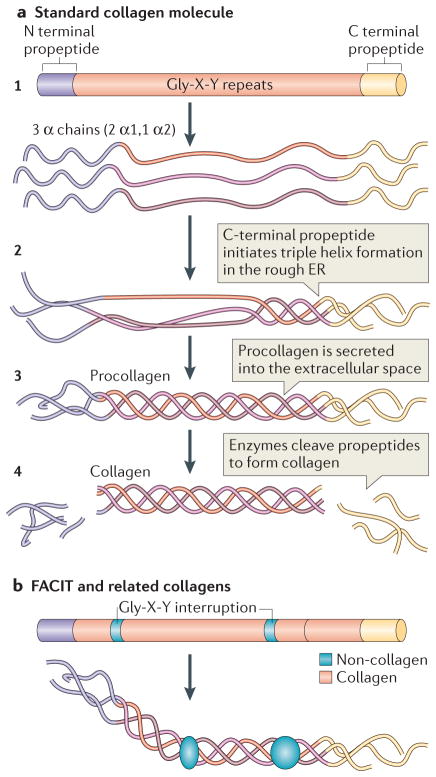
a | The standard fibrillar collagen molecule is characterized by amino- and carboxy-terminal propeptide sequences, which flank a series of Gly-X-Y repeats (where X and Y represent any amino acids but are frequently proline and hydroxyproline) (step 1). These form the central triple helical structure of procollagen and collagen. Three α-chains (the illustration shows two α1-chains and one α2-chain, which is representative of type I collagen) are intracellularly assembled into the triple helix following initiation of this process by the C-terminal domain (step 2). Procollagen is secreted by cells into the extracellular space (step 3) and converted into collagen by the removal of the N- and C-propeptides via metalloproteinase enzymes (step 4). b | Fibril-associated collagens with interrupted triple helices (FACIT) and related collagens have a different structure to standard fibrillar collagen; they contain non-collagenous regions — that is, non-triple helical sequences. These lead to kinks in the resulting macromolecular structure that straighten under small strains. Figure part a is modified, with permission, from REF © 2012 Fan et al.; licensee BioMed Central Ltd, and from REF. , Klug, William S.; Cummings, Michael R., Concepts of Genetics, 5th Edition, © 1997. Reprinted by permission of Pearson Education, Inc., Upper Saddle River, NJ. Figure part b: this figure was originally published in Biochem. J. Jäälinoja, J., Ylöstalo, J., Beckett, W., Hulmes, D. J. S. & Ala-Kokko, L., Trimerization of collagen IX alpha-chains does not require the presence of the COL1 and NC1 domains. Biochem. J. 2008; 409: 545–554 © the Biochemical Society.
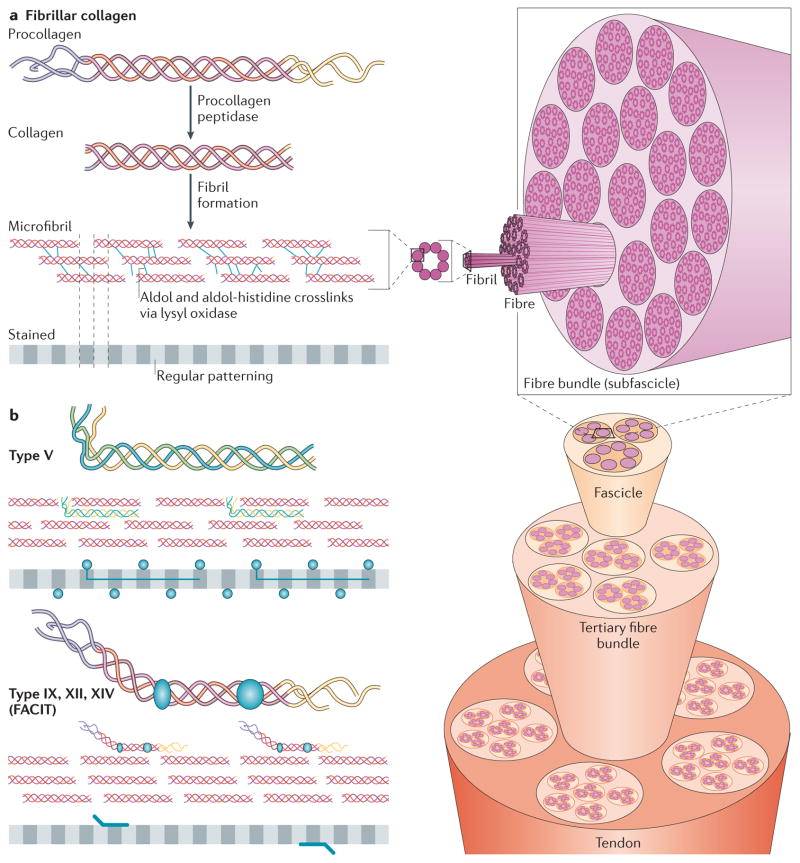
a | After procollagen is secreted into the extracellular space, collagen type-specific metalloproteinase enzymes remove the amino- and carboxy-propeptides. Collagen is assembled into cross-striated microfibrils that occur in the extracellular matrix of connective tissues. This can be observed via electron microscopy as regularly spaced bands, as represented by the grey, striped bar. The position of non-fibrillar collagens in this structure is shown in similar diagrams in part b. Short microfibrils merge into mature fibrils through longitudinal and axial growth. To form mature fibres, lysyl oxidase catalyses the formation of intramolecular and intermolecular covalent crosslinks between collagen molecules. Fibres are bundled together within a connective tissue and stabilized via interactions with fibril-associated collagens with interrupted triple helices (FACIT) collagens and small leucine-rich repeat proteoglycans (SLRPs) into linear structures capable of transmitting tensile forces. Specifically in the case of tendons, fibres ranging in size from 1 μm to 20 μm are bundled into 15 400 μm diameter triangular subfascicles, subfascicles are bundled into 150 1000 μm fascicles and fascicles are grouped into tertiary bundles ranging from 1000 μm to 3000 μm. Finally, multiple tertiary bundles are enclosed by the epitenon connective tissue, which contains the vascular, lymphatic and nerve sources for the tendon tissue. b | In addition to fibrillar collagen, other collagen subtypes, such as type V and FACIT collagens, are incorporated into the fibril structure. Type V collagen is inserted between strands of the microfibril, and FACIT collagens cling to the surface of the microfibril and work with SLRPs to stabilize higher order structures. Figure part a is modified, with permission, from REF. © 2012 Fan et al.; licensee BioMed Central Ltd, from Klug, William S.; Cummings, Michael R., Concepts of Genetics, 5th Edition, © 1997 (REF. 136). Reprinted by permission of Pearson Education, Inc., Upper Saddle River, NJ, and from REF. , reprinted from Trends Biotechnol., 26, Liu, Y., Ramanath, H. S. & Wang, D.-A., Tendon tissue engineering using scaffold enhancing strategies, 201–209, Copyright (2008), with permission from Elsevier. Figure part b: this figure was originally published in Biochem. J. Jäälinoja, J., Ylöstalo, J., Beckett, W., Hulmes, D. J. S. & Ala-Kokko, L., Trimerization of collagen IX alpha-chains does not require the presence of the COL1 and NC1 domains. Biochem. J. 2008; 409: 545–554 © the Biochemical Society.
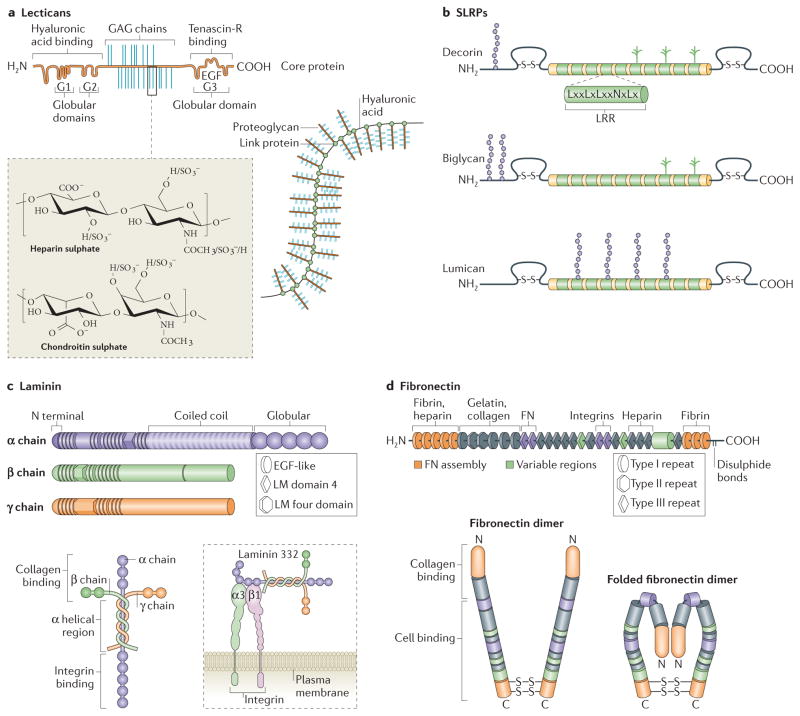
Lecticans (aggrecan is shown; part a) and small leucine-rich repeat proteoglycans (SLRPs; decorin, biglycan and lumican are shown; part b) are major proteoglycans (PGs). The SLRPs are characterized by differences in their amino terminus structure and highly conserved leucine rich repeats (LRR) in the core molecule. Lecticans have a core protein with binding domains for glycosaminoglycan (GAG) chains flanked by globular domains that interact with hyaluronic acid (at the N terminus) and tenascin R (at the carboxy terminus). Common GAGs include chondroitin sulphate and heparan sulphate, the chemical structures of which are depicted. The assembled bottle-brush-like aggrecan link protein hyaluronic acid structure is shown. Laminins are formed by the incorporation of α, β and γ chains into a cruciform, Y-shaped or rod-like structure (part c). These chains are characterized by different domains, as shown. The domain structures depicted represent only one isoform for each chain type, but major differences between the basic composition of the chains (such as the lack of globular regions in β and γ chains) are also present in the other isoforms. Laminins interact with cell surface receptors, such as integrins, primarily through globular domains in the chain. Fibronectin domain structure and the domains to which extracellular matrix (ECM) molecules and cell surface receptors bind are indicated (part d). The fibronectin molecule forms a dimer through disulphide bonds on its C terminus. The folded fibronectin molecule forms via ionic interactions between type III domains of neighbouring molecules and is deformed by mechanical force to reveal cryptic binding sites for other fibronectin molecules and cell surface receptors when interacting with cells. Figure part a: this image originally appeared in Diamond Light Source Ltd’s Annual Report 2010 (p72–73) and has been reproduced with permission from Diamond Light Source Ltd, the University of Liverpool, T. R. Rudd, M. A. Skidmore, S. E. Guimond, R. Xu, R. Hussain, D. G. Fernig, G. Siligardi and E. A. Yates http://www.diamond.ac.uk/Science/Research/Highlights/study2.html . Figure part b is modified from REF. , Nature Publishing Group. Figure part c is modified, with permission, from Yurchenco, P. D. Basement membranes: cell scaffoldings and signaling platforms. Cold Spring Harb. Perspect. Biol.
3, a004911 (2011), Cold Spring Harbour Laboratory Press, and REF. , Nature Publishing Group. Figure part d is modified, with permission, from Schwarzbauer, J. E. & DeSimone, D. W. Fibronectins, their fibrillogenesis, and in vivo functions. Cold Spring Harb. Perspect. Biol.
3, a005041 (2011), Cold Spring Harbour Laboratory Press, and REF. , modified with permission from Annual Review of Cell Dev. Biol., Volume 26 by Annual Reviews, http://www.annualreviews.org . EGF, epidermal growth factor; LM domain, laminin domain.
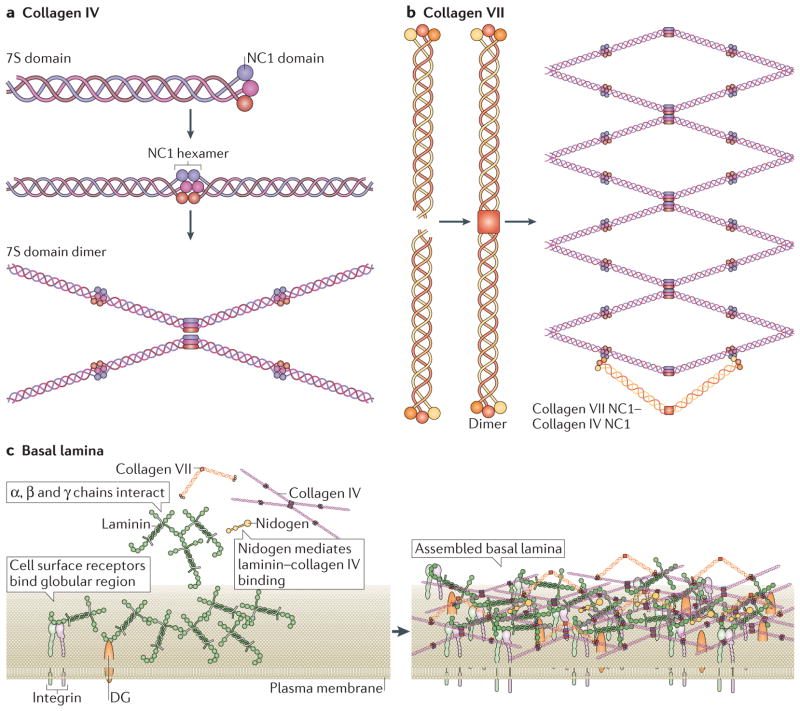
Two major collagens participate in the assembly of the basal lamina: collagen IV and collagen VII. a | Collagen IV is characterized by an amino-terminal 7S domain and a carboxy-terminal NC1 domain these are globular domains that facilitate the formation of hexamers and dimers, respectively. These then form highly ordered lattice structures, as shown. b | Collagen VII forms long dimers that then bend and connect with the globular regions of collagen IV lattices. c | Components of the basal lamina, and how they interact, are shown on the left-hand side. Within the basal lamina, laminins form a sheet on the cell surface by recruiting soluble laminin. This sheet is reinforced via interactions among chains of different laminin molecules and by binding between laminin globular regions and cell surface receptors (for example, integrins and dystroglycan). Collagen IV binds to laminins via nidogens and forms a scaffold on top of the laminins. This scaffold is further secured by collagen VII, which, in its unbent form (sometimes termed anchoring fibrils), can link the basal lamina to the reticular lamina to form the basement membrane. Assembled basal lamina is shown on the right-hand side. Figure parts a and b are modified from REF. , Nature Publishing Group. Figure part c is modified from REF. , Nature Publishing Group, with permission, from Yurchenco, P. D. Basement membranes: cell scaffoldings and signaling platforms. Cold Spring Harb. Perspect. Biol.
3, a004911 (2011), Cold Spring Harbor Laboratory Press and from REF. , Nature Publishing Group. DG, dystroglycan.
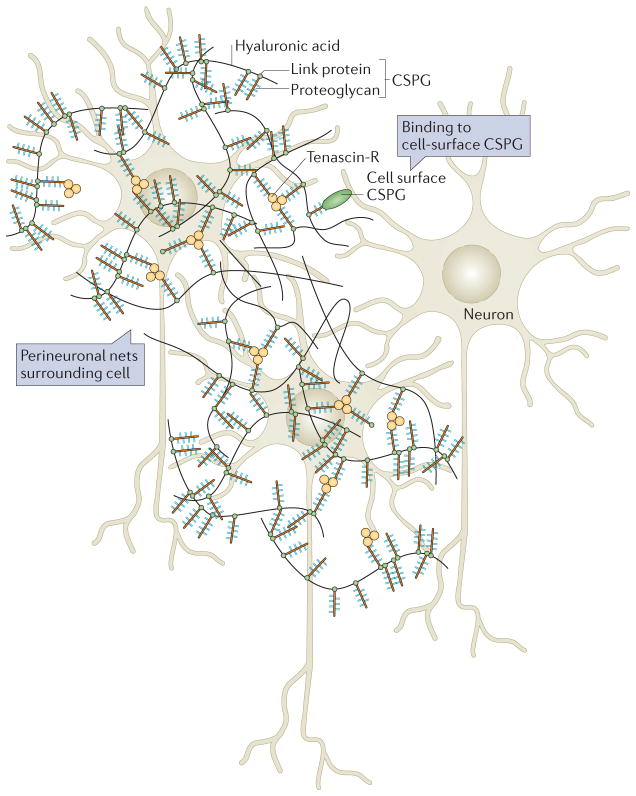
Perineuronal nets (PNNs) surround cells of the brain and act as inhibitors of both growth and migration. PNNs are lattices of hyaluronic acid, proteoglycans and tenascin molecules. The basic structure of a PNN is depicted here. Hyaluronic acid chains coated with proteoglycans are linked by trimeric tenascin R molecules to form a fairly organized structure around neurons. The chondroitin sulphate proteoglycans (CSPGs) within the net bind to cell surface CSPGs and can thus control cell behaviour. Figure modified from REF. , Nature Publishing Group.
Similar articles
-
Targeted proteomics effectively quantifies differences between native lung and detergent-decellularized lung extracellular matrices.Acta Biomater. 2016 Dec;46:91-100. doi: 10.1016/j.actbio.2016.09.043. Epub 2016 Sep 29. Acta Biomater. 2016. PMID: 27693690 Free PMC article.
-
The bi-functional organization of human basement membranes.PLoS One. 2013 Jul 3;8(7):e67660. doi: 10.1371/journal.pone.0067660. Print 2013. PLoS One. 2013. PMID: 23844050 Free PMC article.
-
Mechanical properties of basement membrane in health and disease.Matrix Biol. 2017 Jan;57-58:366-373. doi: 10.1016/j.matbio.2016.07.001. Epub 2016 Jul 17. Matrix Biol. 2017. PMID: 27435904 Review.
-
Extracellular matrix components in the pathogenesis of type 1 diabetes.Curr Diab Rep. 2014 Dec;14(12):552. doi: 10.1007/s11892-014-0552-7. Curr Diab Rep. 2014. PMID: 25344787 Free PMC article. Review.
-
[Structural elements and conformation of basement membranes].Postepy Hig Med Dosw. 2000;54(1):83-97. Postepy Hig Med Dosw. 2000. PMID: 10803026 Review. Polish.
Cited by
-
Design of Bio-Conjugated Hydrogels for Regenerative Medicine Applications: From Polymer Scaffold to Biomolecule Choice.Molecules. 2020 Sep 7;25(18):4090. doi: 10.3390/molecules25184090. Molecules. 2020. PMID: 32906772 Free PMC article. Review.
-
ADAMTS9-Mediated Extracellular Matrix Dynamics Regulates Umbilical Cord Vascular Smooth Muscle Differentiation and Rotation.Cell Rep. 2015 Jun 16;11(10):1519-28. doi: 10.1016/j.celrep.2015.05.005. Epub 2015 May 28. Cell Rep. 2015. PMID: 26027930 Free PMC article.
-
Formation of pre-metastatic bone niche in prostate cancer and regulation of traditional chinese medicine.Front Pharmacol. 2022 Aug 17;13:897942. doi: 10.3389/fphar.2022.897942. eCollection 2022. Front Pharmacol. 2022. PMID: 36059977 Free PMC article. Review.
-
Modulating barriers of tumor microenvironment through nanocarrier systems for improved cancer immunotherapy: a review of current status and future perspective.Drug Deliv. 2020 Dec;27(1):1248-1262. doi: 10.1080/10717544.2020.1809559. Drug Deliv. 2020. PMID: 32865029 Free PMC article. Review.
-
Decellularized extracellular matrix-based disease models for drug screening.Mater Today Bio. 2024 Sep 27;29:101280. doi: 10.1016/j.mtbio.2024.101280. eCollection 2024 Dec. Mater Today Bio. 2024. PMID: 39399243 Free PMC article. Review.
References
-
- Mecham RP. Overview of extracellular matrix. Curr Protoc Cell Biol. 2012;10(Unit 10.1) - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
