

Autocatalytic processing of m-AAA protease subunits in mitochondria
- PMID: 19656850
- PMCID: PMC2754935
- DOI: 10.1091/mbc.e09-03-0218
Autocatalytic processing of m-AAA protease subunits in mitochondria
Abstract
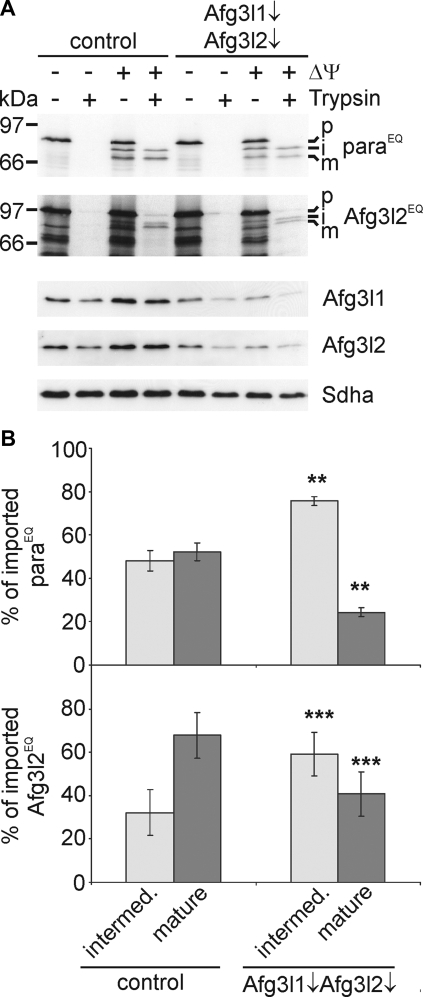
m-AAA proteases are ATP-dependent proteolytic machines in the inner membrane of mitochondria which are crucial for the maintenance of mitochondrial activities. Conserved nuclear-encoded subunits, termed paraplegin, Afg3l1, and Afg3l2, form various isoenzymes differing in their subunit composition in mammalian mitochondria. Mutations in different m-AAA protease subunits are associated with distinct neuronal disorders in human. However, the biogenesis of m-AAA protease complexes or of individual subunits is only poorly understood. Here, we have examined the processing of nuclear-encoded m-AAA protease subunits upon import into mitochondria and demonstrate autocatalytic processing of Afg3l1 and Afg3l2. The mitochondrial processing peptidase MPP generates an intermediate form of Afg3l2 that is matured autocatalytically. Afg3l1 or Afg3l2 are also required for maturation of newly imported paraplegin subunits after their cleavage by MPP. Our results establish that mammalian m-AAA proteases can act as processing enzymes in vivo and reveal overlapping activities of Afg3l1 and Afg3l2. These findings might be of relevance for the pathogenesis of neurodegenerative disorders associated with mutations in different m-AAA protease subunits.
Figures
Similar articles
-
Variable and tissue-specific subunit composition of mitochondrial m-AAA protease complexes linked to hereditary spastic paraplegia.Mol Cell Biol. 2007 Jan;27(2):758-67. doi: 10.1128/MCB.01470-06. Epub 2006 Nov 13. Mol Cell Biol. 2007. PMID: 17101804 Free PMC article.
-
Regulation of OPA1 processing and mitochondrial fusion by m-AAA protease isoenzymes and OMA1.J Cell Biol. 2009 Dec 28;187(7):1023-36. doi: 10.1083/jcb.200906084. J Cell Biol. 2009. PMID: 20038678 Free PMC article.
-
OPA1 processing reconstituted in yeast depends on the subunit composition of the m-AAA protease in mitochondria.Mol Biol Cell. 2007 Sep;18(9):3582-90. doi: 10.1091/mbc.e07-02-0164. Epub 2007 Jul 5. Mol Biol Cell. 2007. PMID: 17615298 Free PMC article.
-
m-AAA proteases, mitochondrial calcium homeostasis and neurodegeneration.Cell Res. 2018 Mar;28(3):296-306. doi: 10.1038/cr.2018.17. Epub 2018 Feb 16. Cell Res. 2018. PMID: 29451229 Free PMC article. Review.
-
ATP-dependent proteases controlling mitochondrial function in the yeast Saccharomyces cerevisiae.Cell Mol Life Sci. 1999 Nov 30;56(9-10):825-42. doi: 10.1007/s000180050029. Cell Mol Life Sci. 1999. PMID: 11212342 Free PMC article. Review.
Cited by
-
Mitochondrial Quality Control Proteases in Neuronal Welfare.J Neuroimmune Pharmacol. 2016 Dec;11(4):629-644. doi: 10.1007/s11481-016-9683-8. Epub 2016 May 2. J Neuroimmune Pharmacol. 2016. PMID: 27137937 Free PMC article. Review.
-
Loss of Mitochondrial AAA Proteases AFG3L2 and YME1L Impairs Mitochondrial Structure and Respiratory Chain Biogenesis.Int J Mol Sci. 2018 Dec 7;19(12):3930. doi: 10.3390/ijms19123930. Int J Mol Sci. 2018. PMID: 30544562 Free PMC article.
-
Early onset and slow progression of SCA28, a rare dominant ataxia in a large four-generation family with a novel AFG3L2 mutation.Eur J Hum Genet. 2010 Aug;18(8):965-8. doi: 10.1038/ejhg.2010.40. Epub 2010 Mar 31. Eur J Hum Genet. 2010. PMID: 20354562 Free PMC article.
-
A snapshot of the Ixodes scapularis degradome.Gene. 2011 Aug 15;482(1-2):78-93. doi: 10.1016/j.gene.2011.04.008. Epub 2011 Apr 28. Gene. 2011. PMID: 21596113 Free PMC article.
-
Multifaceted Roles of AFG3L2, a Mitochondrial ATPase in Relation to Neurological Disorders.Mol Neurobiol. 2024 Jul;61(7):3788-3808. doi: 10.1007/s12035-023-03768-z. Epub 2023 Nov 28. Mol Neurobiol. 2024. PMID: 38012514 Free PMC article. Review.
References
-
- Arlt H., Tauer R., Feldmann H., Neupert W., Langer T. The YTA10–12-complex, an AAA protease with chaperone-like activity in the inner membrane of mitochondria. Cell. 1996;85:875–885. - PubMed
-
- Cagnoli C., et al. Mutations in AFG3L2 gene (SCA28) in autosomal dominant cerebellar ataxias. Annual Meeting of the American Society of Human Genetics,; Philadelphia, PA. 2008.
-
- Casari G., et al. Spastic paraplegia and OXPHOS impairment caused by mutations in paraplegin, a nuclear-encoded mitochondrial metalloprotease. Cell. 1998;93:973–983. - PubMed
-
- Chan D. C. Mitochondria: dynamic organelles in disease, aging, and development. Cell. 2006;125:1241–1252. - PubMed
-
- Cipolat S., et al. Mitochondrial rhomboid PARL regulates cytochrome c release during apoptosis via OPA1-dependent cristae remodeling. Cell. 2006;126:163–175. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
