

doi: 10.1073/pnas.150236297.
Rapid restoration of visual pigment and function with oral retinoid in a mouse model of childhood blindness
Affiliations
- PMID: 10869443
- PMCID: PMC26998
- DOI: 10.1073/pnas.150236297
Item in Clipboard
Rapid restoration of visual pigment and function with oral retinoid in a mouse model of childhood blindness
Proc Natl Acad Sci U S A.
.
Abstract
Mutations in the retinal pigment epithelium gene encoding RPE65 are a cause of the incurable early-onset recessive human retinal degenerations known as Leber congenital amaurosis. Rpe65-deficient mice, a model of Leber congenital amaurosis, have no rod photopigment and severely impaired rod physiology. We analyzed retinoid flow in this model and then intervened by using oral 9-cis-retinal, attempting to bypass the biochemical block caused by the genetic abnormality. Within 48 h, there was formation of rod photopigment and dramatic improvement in rod physiology, thus demonstrating that mechanism-based pharmacological intervention has the potential to restore vision in otherwise incurable genetic retinal degenerations.
Figures
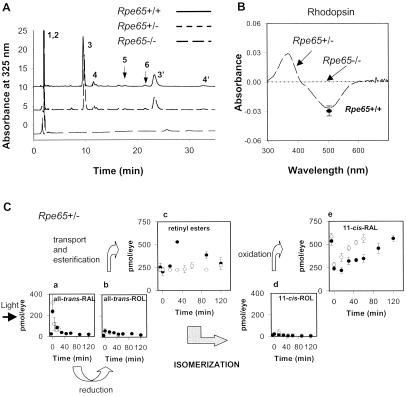
Retinoid and rhodopsin analysis in Rpe65 mice.
(A) Chromatogram of retinoids extracted from eyes of
Rpe65+/+, Rpe65+/−, and
Rpe65−/− mice. The extraction procedures and
derivatization with hydroxylamine to improve quantitative extraction of
retinaldehydes were described previously (see ref. 13). Peaks are
represented as follows: (1, 2) all-trans-retinyl and
11-cis-retinyl esters (truncated to fit in scale);
(3,3′) anti- and syn- of
11-cis-retinal oximes; (4,4′) anti- and
syn- of all-trans-retinal oximes; (5)
11-cis-retinol, and (6)
all-trans-retinol. (B) Difference
spectrum of bleached rhodopsin and rhodopsin extracted from eyes of
Rpe65+/− and Rpe65−/− mice. The
level of rhodopsin in Rpe65+/− mice is within the
range of rhodopsin found in control Rpe65+/+ mice
(●) and undetectable in
Rpe65−/− mice. (C) Aberrant kinetics
of retinoid and rhodopsin recovery in Rpe65+/− mice
after a flash. Photolyzed rhodopsin releases
all-trans-retinal (a), which is reduced
to all-trans-retinol (b), and then
transported to the RPE and esterified to retinyl esters
(c). All-trans-retinol, or its
derivative, is isomerized to 11-cis-retinol
(d), which in turn, is oxidized to
11-cis-retinal (e). The rates of retinoid
formations are compared to the rate obtained for
Rpe65+/+ mice ( ).
).
).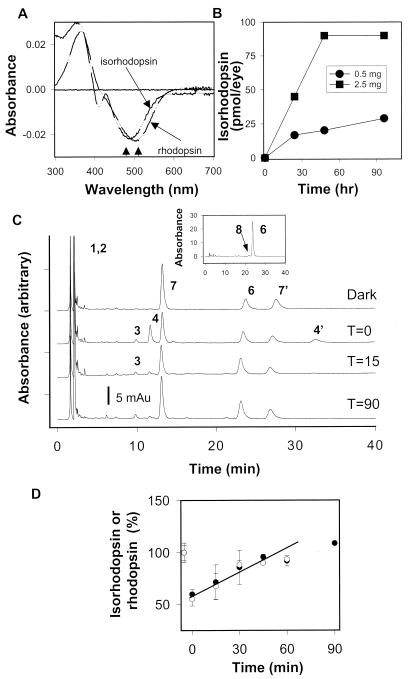
Formation of isorhodopsin and kinetics of the retinoid flow in
Rpe65−/− mice 48 h after
9-cis-retinal gavage. (A) Comparison
spectra of rhodopsin from Rpe65+/+ mice and
isorhodopsin from Rpe65−/− mice 48 h after
9-cis-retinal gavage (2.5 mg). Arrows denote differences
in the absorption maximum for rhodopsin and isorhodopsin.
(B) Isorhodopsin formation in
Rpe65−/− mice at different time points after 0.5 mg
or 2.5 mg of 9-cis-retinal gavage (n
= 2). (C) Chromatograms illustrate the dark recovery of
retinoids in Rpe65−/− mice 48 h after
9-cis-retinal gavage (2.5 mg) after a flash that
bleached ≈45% of isorhodopsin. (C,
Inset) isomeric composition of retinyl esters (8,
9-cis-retinol). (D) Dark recovery of
isorhodopsin in Rpe65−/− mice 48 h after
9-cis-retinal gavage after a flash. For comparison, dark
recovery of rhodopsin is shown for Rpe65+/+ (open
hexagons) (7, 7′). anti- and syn- of
9-cis-Retinal oximes; all other peaks are as in the Fig.
1 legend.
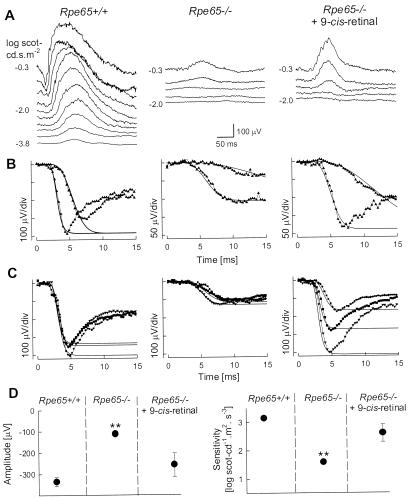
Restoration of retinal function in Rpe65−/−
mice 48 h after 9-cis-retinal gavage.
(A and B) Serial ERG recordings in an
Rpe65−/− mouse before and 48 h after
9-cis-retinal gavage compared to a representative
Rpe65+/+ mouse. (A) Dark-adapted ERGs
to increasing intensities of blue light stimuli (shown to the left of
the traces) in an Rpe65−/− mouse show an elevated
b-wave threshold compared to Rpe65+/+. The same
stimuli after 9-cis-retinal treatment elicit ERGs at a
lower threshold and with larger amplitude b-waves. (B)
Leading edges (initial 4–15 ms depending on response) of dark-adapted
ERG photoresponses (symbols) evoked by 3.6 and 2.2 log
scot-cd⋅s⋅m−2 flashes are fit with a model of
phototransduction (smooth lines). The amplitude and sensitivity of the
Rpe65−/− mouse photoresponses are reduced. After
9-cis-retinal treatment, photoresponses have larger
amplitude and higher sensitivity. (C) Photoresponses in
three Rpe65+/+ mice are compared to an untreated group
of Rpe65−/− mice and a treated group of
Rpe65−/− mice. Lines are the model of rod
phototransduction activation fitted to a pair of photoresponses; only
maximal responses are shown for clarity. (D) Maximum
amplitude and sensitivity parameters of dark-adapted photoresponses in
untreated and treated (48 h after 9-cis-retinal gavage)
Rpe65−/− mice compared to the results in
Rpe65+/+ mice. Untreated animals have significant
differences (**, P < 0.05) in both
parameters when compared to Rpe65+/+ or treated
Rpe65−/− mice. Error bars represent 1 SEM and are
smaller than symbols for some data.
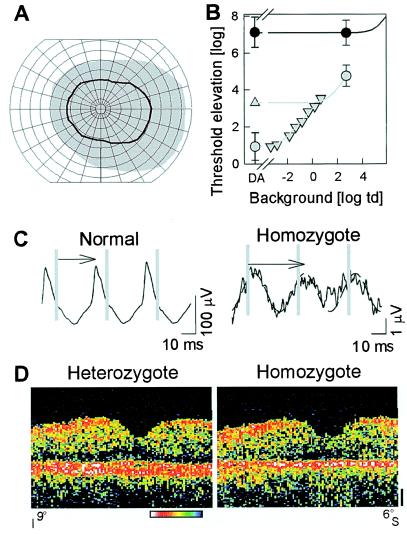
Human phenotype of putative null mutation in RPE65.
(A) Kinetic visual fields with a V-4e test target in the
homozygote (black line) compared to normal (gray region); concentric
circles are at 10o intervals and meridians are at
15o. (B) White stimulus thresholds of the
homozygote (●) shows >6 log units of elevation
under dark-adapted (DA) and >2 log units of elevation under
light-adapted (2.7 log troland) conditions. Normal thresholds (gray
symbols) on increasing background intensities (▿) and during the cone
plateau following a full bleach (▵) are fit with empirical
models of background adaptation (gray lines) to define the rod- and
cone-mediated limbs. The normal cone adaptation model was shifted by
3.8 log units to the right and up to fit the patient data (solid line).
(C) Cone flicker ERGs in homozygote are reduced
≈100-fold in amplitude and abnormally delayed in timing (arrows).
Vertical gray lines mark stimulus times. A 29-Hz sinusoid was fitted to
the homozygote data (dashed lines) to estimate amplitude and timing of
the small signal. (D) Vertical OCT cross-sectional
retinal images through the fovea (9o inferior to
6o superior retina) show central retinal structure is
generally intact in the homozygote (compared to a heterozygous sibling)
except for thinning of the outer retina-choroid complex, the
red/white band toward bottom of panel. Optical reflectivity of
retinal tissue is shown on a logarithmic pseudocolor scale: red and
white are high reflectivity and blue and black are low reflectivity.
Vitreous is toward the top and sclera is toward the bottom.
Similar articles
-
Recovery of visual functions in a mouse model of Leber congenital amaurosis.J Biol Chem. 2002 May 24;277(21):19173-82. doi: 10.1074/jbc.M112384200. Epub 2002 Mar 15. J Biol Chem. 2002. PMID: 11897783 Free PMC article.
-
Retinal degeneration 12 (rd12): a new, spontaneously arising mouse model for human Leber congenital amaurosis (LCA).Mol Vis. 2005 Feb 28;11:152-62. Mol Vis. 2005. PMID: 15765048
-
Rpe65-/- and Lrat-/- mice: comparable models of leber congenital amaurosis.Invest Ophthalmol Vis Sci. 2008 Jun;49(6):2384-9. doi: 10.1167/iovs.08-1727. Epub 2008 Feb 22. Invest Ophthalmol Vis Sci. 2008. PMID: 18296659 Free PMC article.
-
RPE65: role in the visual cycle, human retinal disease, and gene therapy.Ophthalmic Genet. 2009 Jun;30(2):57-62. doi: 10.1080/13816810802626399. Ophthalmic Genet. 2009. PMID: 19373675 Free PMC article. Review.
-
Leber's congenital amaurosis and RPE65.Int Ophthalmol Clin. 2001 Winter;41(1):73-82. doi: 10.1097/00004397-200101000-00008. Int Ophthalmol Clin. 2001. PMID: 11198148 Review. No abstract available.
Cited by
-
M-opsin protein degradation is inhibited by MG-132 in Rpe65⁻/⁻ retinal explant culture.Mol Vis. 2012;18:1516-25. Epub 2012 Jun 13. Mol Vis. 2012. PMID: 22736942 Free PMC article.
-
A point mutation in Semaphorin 4A associates with defective endosomal sorting and causes retinal degeneration.Nat Commun. 2013;4:1406. doi: 10.1038/ncomms2420. Nat Commun. 2013. PMID: 23360997 Free PMC article.
-
Chemical chaperone TUDCA preserves cone photoreceptors in a mouse model of Leber congenital amaurosis.Invest Ophthalmol Vis Sci. 2012 Jun 5;53(7):3349-56. doi: 10.1167/iovs.12-9851. Invest Ophthalmol Vis Sci. 2012. PMID: 22531707 Free PMC article.
-
Molecular Basis for Vitamin A Uptake and Storage in Vertebrates.Nutrients. 2016 Oct 26;8(11):676. doi: 10.3390/nu8110676. Nutrients. 2016. PMID: 27792183 Free PMC article. Review.
-
The Rpe65 Leu450Met variation increases retinal resistance against light-induced degeneration by slowing rhodopsin regeneration.J Neurosci. 2001 Jan 1;21(1):53-8. doi: 10.1523/JNEUROSCI.21-01-00053.2001. J Neurosci. 2001. PMID: 11150319 Free PMC article.
References
-
- Polans A, Baehr W, Palczewski K. Trends Neurosci. 1996;19:547–554. - PubMed
-
- Lagnado L, Baylor D. Neuron. 1992;8:995–1002. - PubMed
-
- Rando R R. Chem Biol. 1996;3:255–262. - PubMed
-
- Palczewski K, Saari J C. Curr Opin Neurobiol. 1997;7:500–504. - PubMed
-
- Bavik C O, Busch C, Eriksson U. J Biol Chem. 1992;267:23035–23042. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
