

The Borrelia burgdorferi Adenylate Cyclase, CyaB, Is Important for Virulence Factor Production and Mammalian Infection
- PMID: 34113333
- PMCID: PMC8186283
- DOI: 10.3389/fmicb.2021.676192
The Borrelia burgdorferi Adenylate Cyclase, CyaB, Is Important for Virulence Factor Production and Mammalian Infection
Abstract
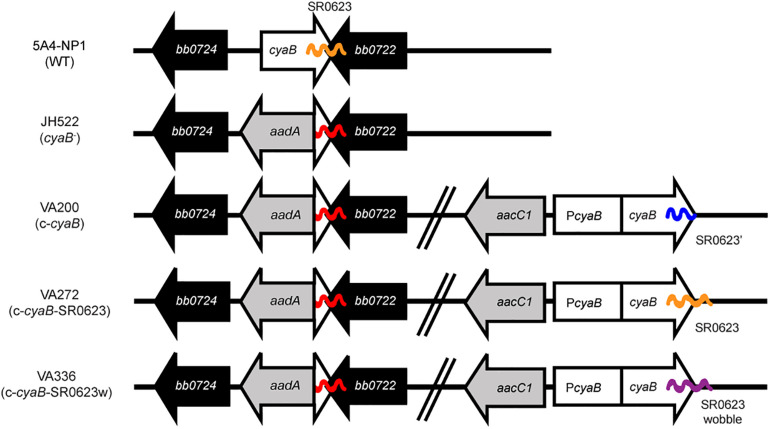
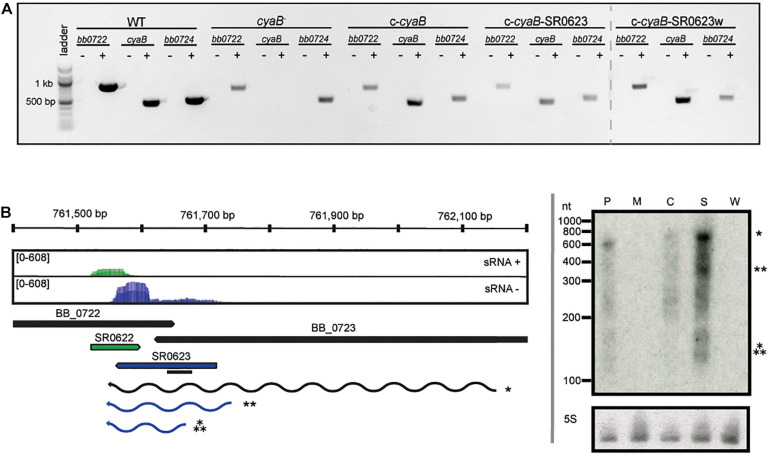
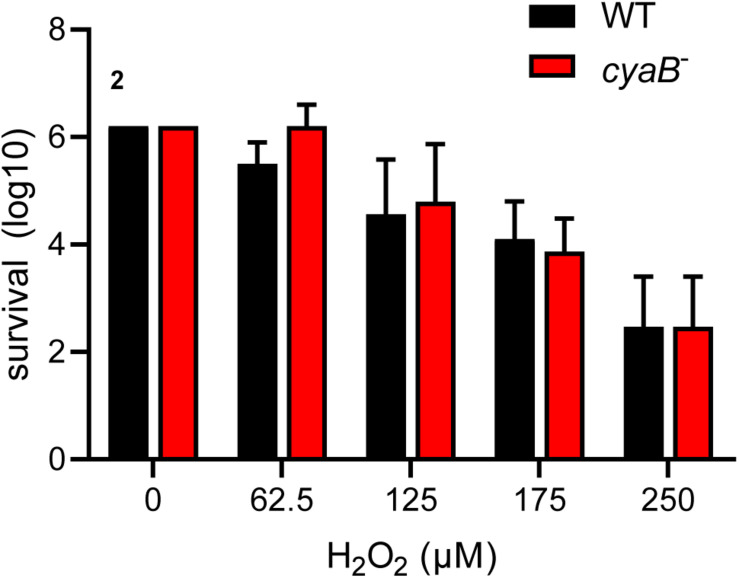
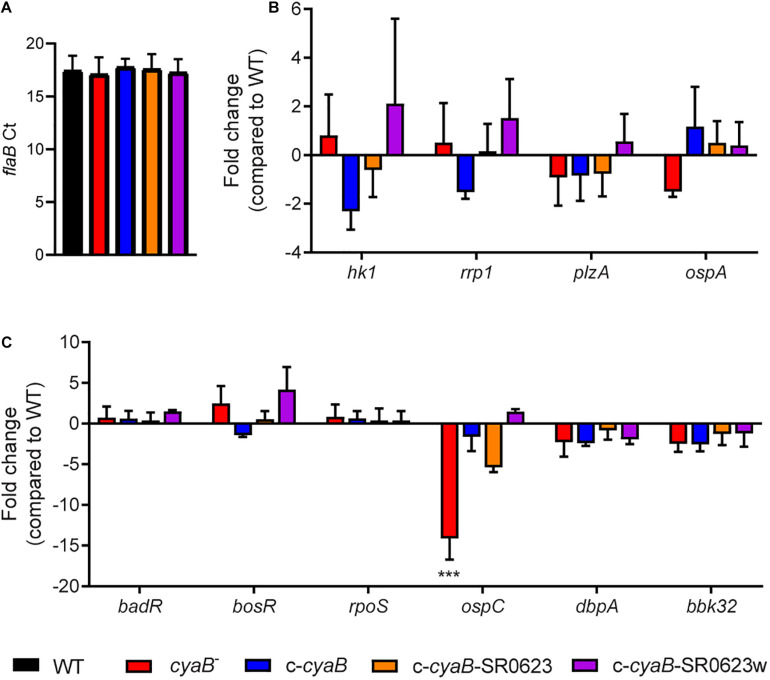
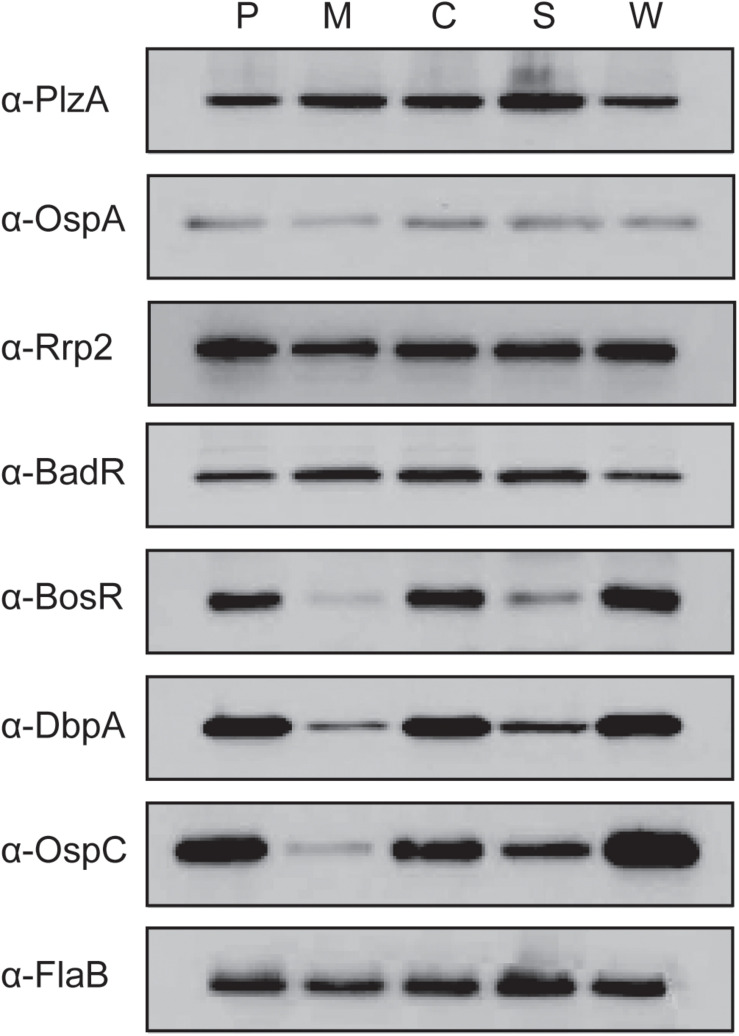
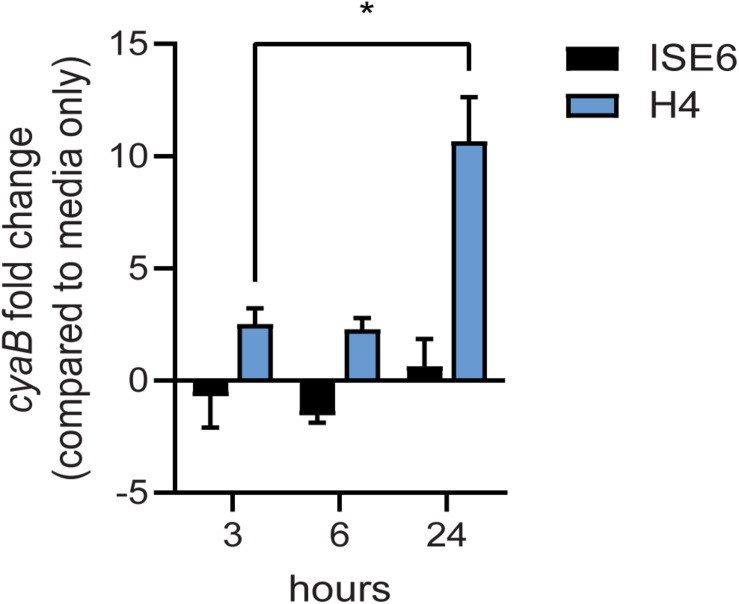
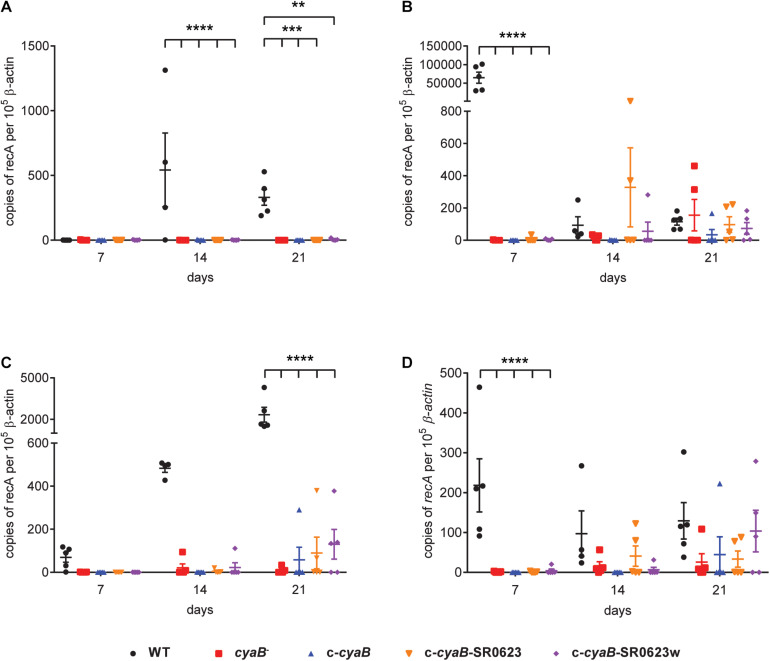
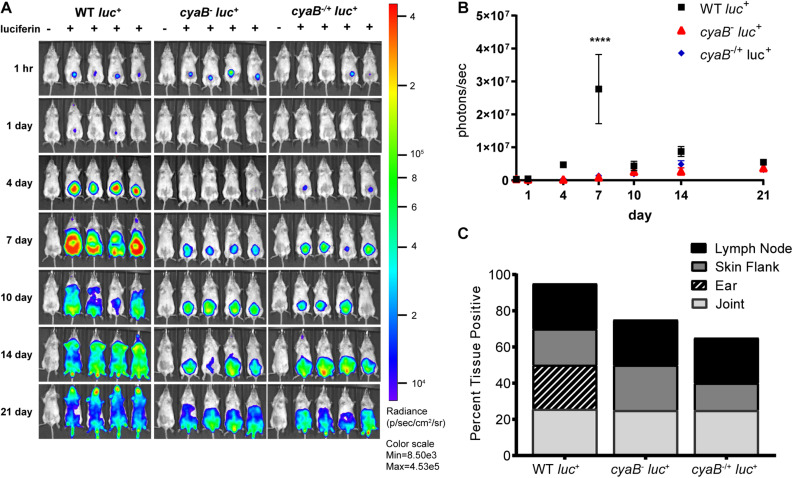
Borrelia burgdorferi, the causative agent of Lyme disease, traverses through vastly distinct environments between the tick vector and the multiple phases of the mammalian infection that requires genetic adaptation for the progression of pathogenesis. Borrelial gene expression is highly responsive to changes in specific environmental signals that initiate the RpoS regulon for mammalian adaptation, but the mechanism(s) for direct detection of environmental cues has yet to be identified. Secondary messenger cyclic adenosine monophosphate (cAMP) produced by adenylate cyclase is responsive to environmental signals, such as carbon source and pH, in many bacterial pathogens to promote virulence by altering gene regulation. B. burgdorferi encodes a single non-toxin class IV adenylate cyclase (bb0723, cyaB). This study investigates cyaB expression along with its influence on borrelial virulence regulation and mammalian infectivity. Expression of cyaB was specifically induced with co-incubation of mammalian host cells that was not observed with cultivated tick cells suggesting that cyaB expression is influenced by cellular factor(s) unique to mammalian cell lines. The 3' end of cyaB also encodes a small RNA, SR0623, in the same orientation that overlaps with bb0722. The differential processing of cyaB and SR0623 transcripts may alter the ability to influence function in the form of virulence determinant regulation and infectivity. Two independent cyaB deletion B31 strains were generated in 5A4-NP1 and ML23 backgrounds and complemented with the cyaB ORF alone that truncates SR0623, cyaB with intact SR0623, or cyaB with a mutagenized full-length SR0623 to evaluate the influence on transcriptional and posttranscriptional regulation of borrelial virulence factors and infectivity. In the absence of cyaB, the expression and production of ospC was significantly reduced, while the protein levels for BosR and DbpA were substantially lower than parental strains. Infectivity studies with both independent cyaB mutants demonstrated an attenuated phenotype with reduced colonization of tissues during early disseminated infection. This work suggests that B. burgdorferi utilizes cyaB and potentially cAMP as a regulatory pathway to modulate borrelial gene expression and protein production to promote borrelial virulence and dissemination in the mammalian host.
Keywords: Borrelia burgdorferi spirochete; CyaB; Lyme disease; adenylate cyclase (AC); cyclic nucleotides; secondary messenger; small RNA (sRNA).
Copyright © 2021 Ante, Farris, Saputra, Hall, O’Bier, Oliva Chávez, Marconi, Lybecker and Hyde.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Phosphoenolpyruvate Phosphotransferase System Components Modulate Gene Transcription and Virulence of Borrelia burgdorferi.Infect Immun. 2015 Dec 28;84(3):754-64. doi: 10.1128/IAI.00917-15. Infect Immun. 2015. PMID: 26712207 Free PMC article.
-
RpoS is not central to the general stress response in Borrelia burgdorferi but does control expression of one or more essential virulence determinants.Infect Immun. 2004 Nov;72(11):6433-45. doi: 10.1128/IAI.72.11.6433-6445.2004. Infect Immun. 2004. PMID: 15501774 Free PMC article.
-
Two Distinct Mechanisms Govern RpoS-Mediated Repression of Tick-Phase Genes during Mammalian Host Adaptation by Borrelia burgdorferi, the Lyme Disease Spirochete.mBio. 2017 Aug 22;8(4):e01204-17. doi: 10.1128/mBio.01204-17. mBio. 2017. PMID: 28830947 Free PMC article.
-
Borrelia burgdorferi Keeps Moving and Carries on: A Review of Borrelial Dissemination and Invasion.Front Immunol. 2017 Feb 21;8:114. doi: 10.3389/fimmu.2017.00114. eCollection 2017. Front Immunol. 2017. PMID: 28270812 Free PMC article. Review.
-
Immune evasion of Borrelia burgdorferi: insufficient killing of the pathogens by complement and antibody.Int J Med Microbiol. 2002 Jun;291 Suppl 33:141-6. doi: 10.1016/s1438-4221(02)80027-3. Int J Med Microbiol. 2002. PMID: 12141738 Review.
Cited by
-
Pathogenicity and virulence of Borrelia burgdorferi.Virulence. 2023 Dec;14(1):2265015. doi: 10.1080/21505594.2023.2265015. Epub 2023 Oct 9. Virulence. 2023. PMID: 37814488 Free PMC article. Review.
-
Genomic analysis of Elizabethkingia species from aquatic environments: Evidence for potential clinical transmission.Curr Res Microb Sci. 2021 Nov 26;3:100083. doi: 10.1016/j.crmicr.2021.100083. eCollection 2022. Curr Res Microb Sci. 2021. PMID: 34988536 Free PMC article.
References
-
- Adams P. P., Flores Avile C., Popitsch N., Bilusic I., Schroeder R., Lybecker M., et al. (2017). In vivo expression technology and 5′ end mapping of the borrelia burgdorferi transcriptome identify novel RNAs expressed during mammalian infection. Nucleic Acids Res. 45 775–792. 10.1093/nar/gkw1180 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous