

doi: 10.1128/JVI.02262-14.
Epub 2014 Nov 26.
SAMD9 is an innate antiviral host factor with stress response properties that can be antagonized by poxviruses
Affiliations
- PMID: 25428864
- PMCID: PMC4300762
- DOI: 10.1128/JVI.02262-14
Item in Clipboard
SAMD9 is an innate antiviral host factor with stress response properties that can be antagonized by poxviruses
J Virol.
2015 Feb.
Abstract
We show that SAMD9 is an innate host antiviral stress response element that participates in the formation of antiviral granules. Poxviruses, myxoma virus and vaccinia virus specifically, utilize a virus-encoded host range factor(s), such as a member of the C7L superfamily, to antagonize SAMD9 to prevent granule formation in a eukaryotic initiation factor 2α (eIF2α)-independent manner. When SAMD9 is stimulated due to failure of the viral antagonism during infection, the resulting antiviral granules exhibit properties different from those of the canonical stress granules.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
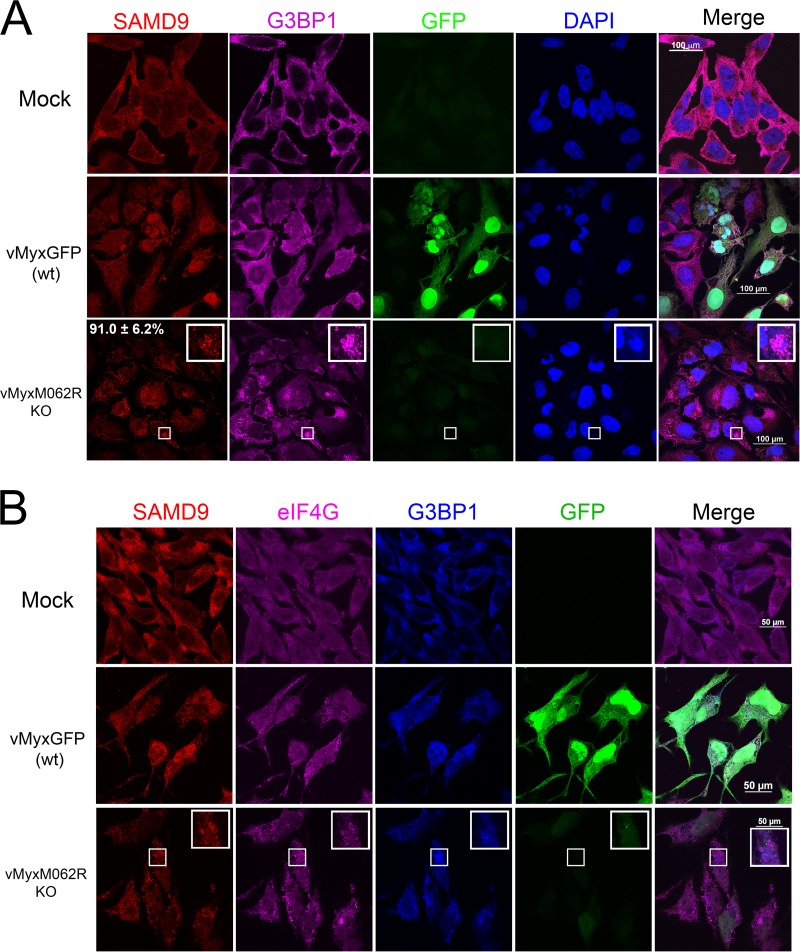
Infection by MYXV-M062R-null (vMyxM062RKO) leads to the formation of SAMD9 granules. (A) SAMD9 forms a granule structure during vMyxM062RKO infection. HeLa cells were mock treated or infected with wild-type MYXV [vMyxGFP (wt)] or vMyxM062RKO at a multiplicity of infection (MOI) of 10 for 18 h. Cells were fixed with 4% paraformaldehyde and permeabilized with cold methanol before they were incubated with primary antibodies (SAMD9 and G3BP1; Sigma-Aldrich and Santa Cruz Biotechnologies, respectively) and then secondary antibodies (Invitrogen). Samples were mounted for fluorescence microscopy (Nikon C2 confocal microscope). The scale bar represents a length of 100 μm. A total of 91.0% (±6.2% SD) of infected cells (GFP positive and/or viral factory positive) showed SAMD9 antiviral granules; within the viral factories, G3BP1 colocalizes with SAMD9 (the quantification was conducted from three independent experiments and four random views of image with 15 to 31 cells per view in each experiment). (B) SAMD9 granules colocalize with markers of stress granules (SGs). HeLa cells were mock treated or infected with wild-type MYXV or vMyxM062RKO viruses at a MOI of 10 for 18 h before immunofluorescence (IF) staining for SAMD9 (Alexa Fluor 594), G3BP1 (Alexa Fluor 350), and eIF4G (Alexa Fluor 647) was performed as described for panel A. Fluorescent images were captured with a Nikon C2 confocal microscope at 100× magnification. Scale bar, 50 μm.
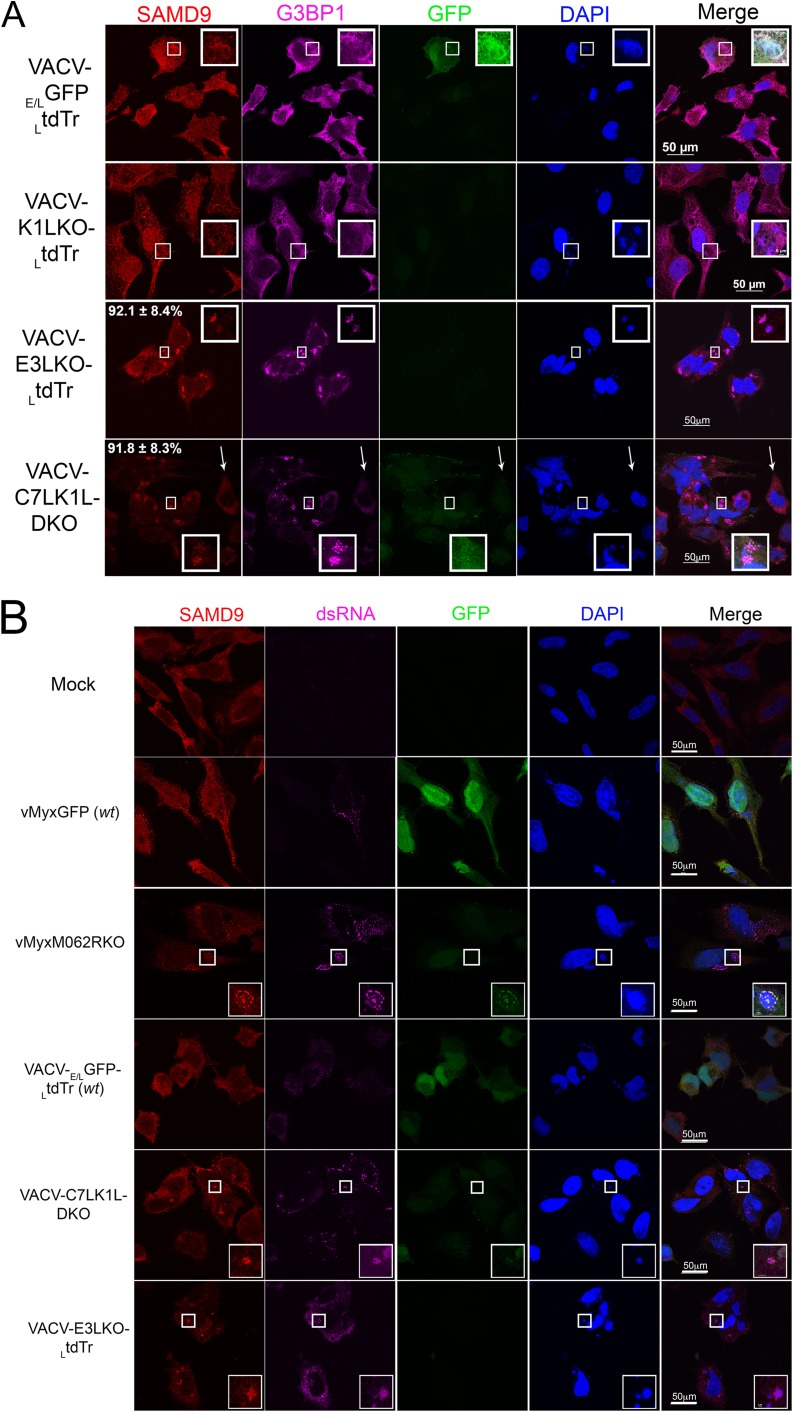
Abortive infection by E3L knockout VACV (VACV-E3LKO-LtdTr) or C7LK1L double-knockout VACV (VACV-C7LK1L-DKO) stimulates SAMD9 granule formation. (A) SAMD9 antiviral granules form after VACV-E3LKO-LtdTr (92.1% ± 8.4% SD) or VACV-C7LK1L-DKO (91.8% ± 8.3% SD) infection; meanwhile, G3BP1 colocalizes with SAMD9 in the viral factories. Wild-type or knockout VACV infection was conducted at a MOI of 5 for 5 h p.i. on HeLa cells before cells were fixed and permeabilized for IF staining of SAMD9 (Alexa Fluor 594) and G3BP1 (Alexa Fluor 647). Nuclei and viral factories were stained with DAPI (4′,6-diamidino-2-phenylindole). Low-level of tdTr expression driven by the late poxvirus promoter is not detectable at this time of the infection, and thus it does not interfere with the observation of Alexa Fluor 594. Fluorescent images were captured with a Nikon C2 confocal microscope. Quantification was conducted from the results of up to five independent experiments. The arrows point to an uninfected cell in the VACV-C7LK1L-DKO infection. Scale bar, 50 μm. (B) Double-stranded RNA can be detected within the SAMD9 granules. Infection by MYXV and VACV was conducted at 19 and 5 h p.i., respectively. After fixing and permeabilization, cells were probed with SAMD9, dsRNA (English and Scientific Consulting Kft.), and DAPI for IF. Fluorescence microscopic observation was performed as described for panel A. Scale bar, 50 μm. (C) The formation of SAMD9 antiviral granules by poxvirus infection is eIF2α phosphorylation independent. HeLa cells were infected with VACV (lane 3, VACV-E/LGFP/LtdTr; lane 4, VACV-K1LKO-LtdTr; lane 5, VACV-C7LK1L-DKO; lane 6, VACV-E3LKO-LtdTr) for 5 h at a MOI of 5 or with MYXV (lane 7, vMyxM062RKO; lane 8, vMyxGFP) for 19 h before they were harvested for Western blotting. As controls, cells were either mock treated (lane 1) or treated with sodium arsenate (SA) (1 mM) for 1 h (lane 2). Total proteins of 20 μg were separated using 12% SDS-PAGE before they were transferred to a polyvinylidene difluoride (PVDF) membrane for probing with phosphorylated eIF2α (Cell Signaling) and later with total eIF2α (Cell Signaling).
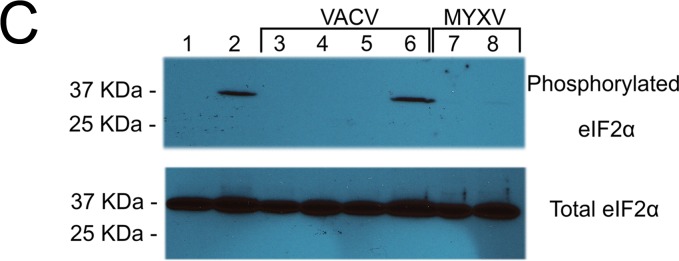
Abortive infection by E3L knockout VACV (VACV-E3LKO-LtdTr) or C7LK1L double-knockout VACV (VACV-C7LK1L-DKO) stimulates SAMD9 granule formation. (A) SAMD9 antiviral granules form after VACV-E3LKO-LtdTr (92.1% ± 8.4% SD) or VACV-C7LK1L-DKO (91.8% ± 8.3% SD) infection; meanwhile, G3BP1 colocalizes with SAMD9 in the viral factories. Wild-type or knockout VACV infection was conducted at a MOI of 5 for 5 h p.i. on HeLa cells before cells were fixed and permeabilized for IF staining of SAMD9 (Alexa Fluor 594) and G3BP1 (Alexa Fluor 647). Nuclei and viral factories were stained with DAPI (4′,6-diamidino-2-phenylindole). Low-level of tdTr expression driven by the late poxvirus promoter is not detectable at this time of the infection, and thus it does not interfere with the observation of Alexa Fluor 594. Fluorescent images were captured with a Nikon C2 confocal microscope. Quantification was conducted from the results of up to five independent experiments. The arrows point to an uninfected cell in the VACV-C7LK1L-DKO infection. Scale bar, 50 μm. (B) Double-stranded RNA can be detected within the SAMD9 granules. Infection by MYXV and VACV was conducted at 19 and 5 h p.i., respectively. After fixing and permeabilization, cells were probed with SAMD9, dsRNA (English and Scientific Consulting Kft.), and DAPI for IF. Fluorescence microscopic observation was performed as described for panel A. Scale bar, 50 μm. (C) The formation of SAMD9 antiviral granules by poxvirus infection is eIF2α phosphorylation independent. HeLa cells were infected with VACV (lane 3, VACV-E/LGFP/LtdTr; lane 4, VACV-K1LKO-LtdTr; lane 5, VACV-C7LK1L-DKO; lane 6, VACV-E3LKO-LtdTr) for 5 h at a MOI of 5 or with MYXV (lane 7, vMyxM062RKO; lane 8, vMyxGFP) for 19 h before they were harvested for Western blotting. As controls, cells were either mock treated (lane 1) or treated with sodium arsenate (SA) (1 mM) for 1 h (lane 2). Total proteins of 20 μg were separated using 12% SDS-PAGE before they were transferred to a polyvinylidene difluoride (PVDF) membrane for probing with phosphorylated eIF2α (Cell Signaling) and later with total eIF2α (Cell Signaling).
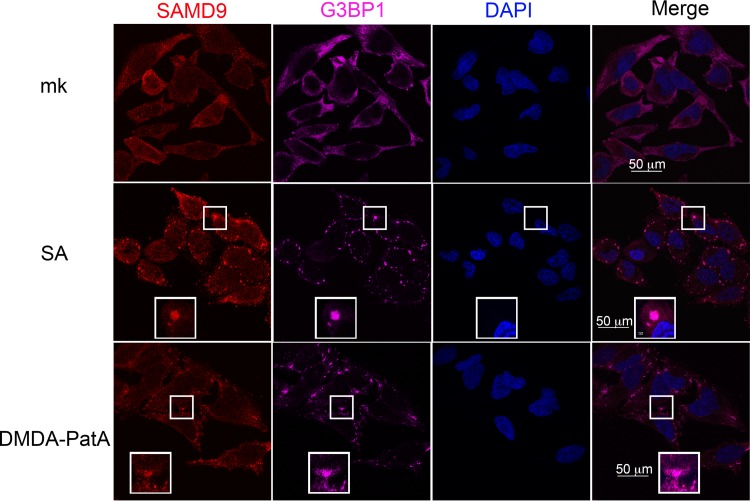
SAMD9 is a stress-responding element that participates in stress granule formation. HeLa cells are treated with sodium arsenate (SA) (1 mM) or DMDA-Pat A (1 μM) for 1 h before IF staining of SAMD9, G3BP1, and nuclei as described for Fig. 1 and 2. mk, mock treatment.

Knocking down SAMD9 rescues MYXV-M062R-null (vMyxM062RKO) and C7LK1L double-knockout VACV (VACV-C7LK1L-DKO) by disengaging the antiviral granule formation. (A) SAMD9 is responsible for the antiviral granule formation stimulated by either vMyxM062RKO (a) or VACV-C7LK1L-DKO (b) but not for the antiviral granule formation stimulated by E3L-knockout VACV (VACV-E3LKO-LtdTr) (c). Antiviral granule formation was investigated at 19 h p.i. for vMyxM062RKO and at 5 h p.i. for VACV infection of normal HeLa cells or SAMD9 knockdown (SAMD9KD) at a MOI of 10 and a MOI of 5, respectively. After IF staining of SAMD9, G3BP1, and nuclei, images were captured with a Nikon c2 confocal microscope. Scale bar, 50 μm. (B) Knocking down SAMD9 expression rescues M062R-null MYXV replication. HeLa cells stably expressing shRNAs targeting SAMD9 (SAMD9KD) showed significantly reduced SAMD9 protein levels compared with control HeLa cells (con). A total protein of 40 μg was separated on SDS-PAGE for Western blotting against SAMD9 and β-actin. Multiple-step growth curve analysis of MYXV-M062R-null was conducted on control HeLa (con) or SAMD9 knockdown (SAMD9KD) HeLa cells. Duplicate of infection at each time point per virus was conducted. Titration was performed on BSC40 cells in triplicate at each dilution. The growth curve shown is one view representative of the results of two independent experiments. ffu, focus-forming units. (C) Knocking down SAMD9 expression rescues viral replication of VACV-C7LK1L-DKO. A549 cells stably expressing control shRNA (A549-con) or shRNAs targeting SAMD9 (A549-SAMD9KD) were engineered by infecting cells with lentivirus containing corresponding shRNAs. Western blot analysis was conducted with a total protein of 40 μg from either A549-con or A549-SAMD9KD by probing against SAMD9 and β-actin. The experiment designed to construct the multiple-step growth curve of VACV-C7LK1L-DKO was performed by infecting control or knockdown cells at a low MOI of 0.1 followed by harvesting at different time points. At each time point, samples from triplicate infections were collected for titration on BSC40 cells. The growth curve shown is a view representative of the results of two independent experiments.
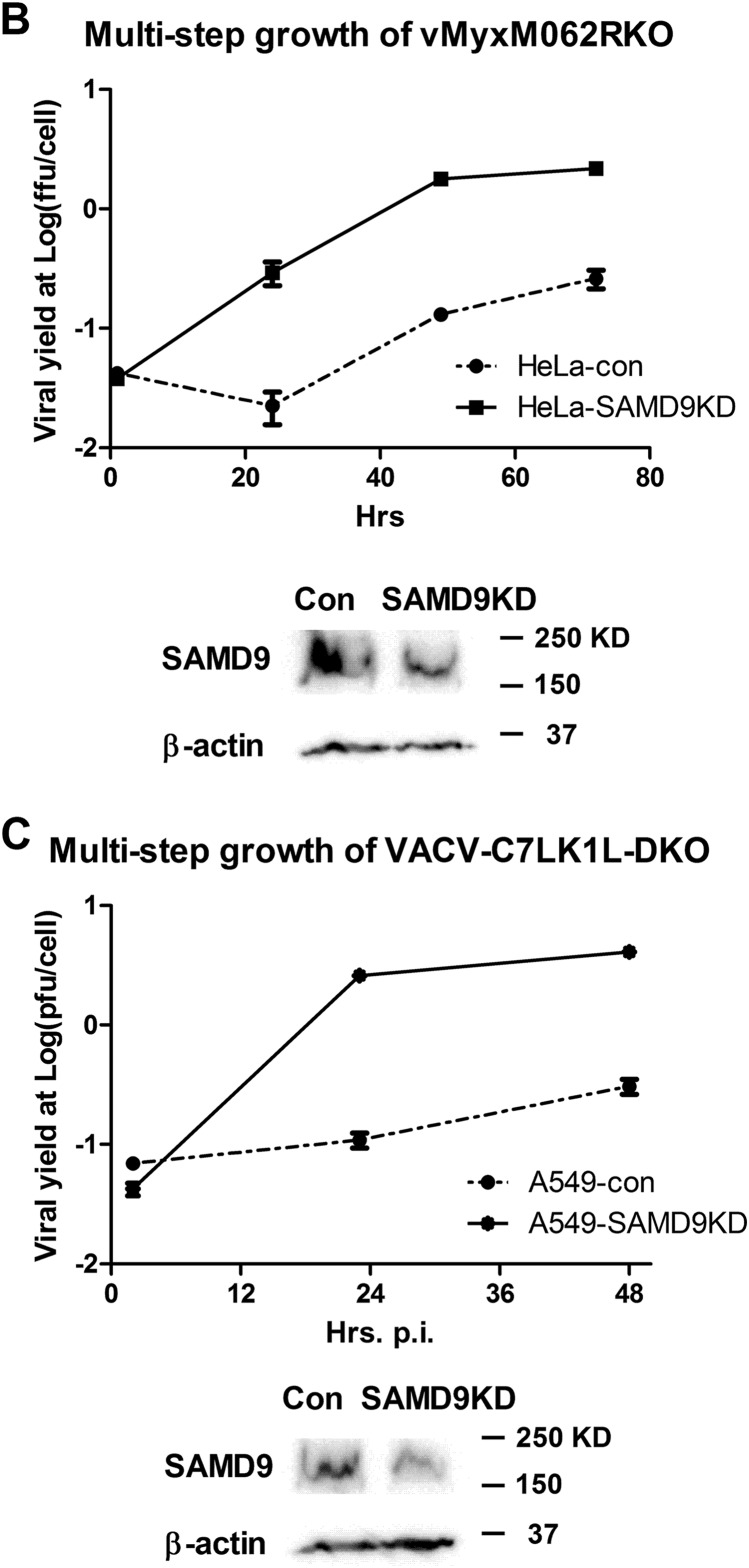
Knocking down SAMD9 rescues MYXV-M062R-null (vMyxM062RKO) and C7LK1L double-knockout VACV (VACV-C7LK1L-DKO) by disengaging the antiviral granule formation. (A) SAMD9 is responsible for the antiviral granule formation stimulated by either vMyxM062RKO (a) or VACV-C7LK1L-DKO (b) but not for the antiviral granule formation stimulated by E3L-knockout VACV (VACV-E3LKO-LtdTr) (c). Antiviral granule formation was investigated at 19 h p.i. for vMyxM062RKO and at 5 h p.i. for VACV infection of normal HeLa cells or SAMD9 knockdown (SAMD9KD) at a MOI of 10 and a MOI of 5, respectively. After IF staining of SAMD9, G3BP1, and nuclei, images were captured with a Nikon c2 confocal microscope. Scale bar, 50 μm. (B) Knocking down SAMD9 expression rescues M062R-null MYXV replication. HeLa cells stably expressing shRNAs targeting SAMD9 (SAMD9KD) showed significantly reduced SAMD9 protein levels compared with control HeLa cells (con). A total protein of 40 μg was separated on SDS-PAGE for Western blotting against SAMD9 and β-actin. Multiple-step growth curve analysis of MYXV-M062R-null was conducted on control HeLa (con) or SAMD9 knockdown (SAMD9KD) HeLa cells. Duplicate of infection at each time point per virus was conducted. Titration was performed on BSC40 cells in triplicate at each dilution. The growth curve shown is one view representative of the results of two independent experiments. ffu, focus-forming units. (C) Knocking down SAMD9 expression rescues viral replication of VACV-C7LK1L-DKO. A549 cells stably expressing control shRNA (A549-con) or shRNAs targeting SAMD9 (A549-SAMD9KD) were engineered by infecting cells with lentivirus containing corresponding shRNAs. Western blot analysis was conducted with a total protein of 40 μg from either A549-con or A549-SAMD9KD by probing against SAMD9 and β-actin. The experiment designed to construct the multiple-step growth curve of VACV-C7LK1L-DKO was performed by infecting control or knockdown cells at a low MOI of 0.1 followed by harvesting at different time points. At each time point, samples from triplicate infections were collected for titration on BSC40 cells. The growth curve shown is a view representative of the results of two independent experiments.
Similar articles
-
Myxoma virus lacking the host range determinant M062 stimulates cGAS-dependent type 1 interferon response and unique transcriptomic changes in human monocytes/macrophages.PLoS Pathog. 2022 Sep 14;18(9):e1010316. doi: 10.1371/journal.ppat.1010316. eCollection 2022 Sep. PLoS Pathog. 2022. PMID: 36103568 Free PMC article.
-
Identification of Restriction Factors by Human Genome-Wide RNA Interference Screening of Viral Host Range Mutants Exemplified by Discovery of SAMD9 and WDR6 as Inhibitors of the Vaccinia Virus K1L-C7L- Mutant.mBio. 2015 Aug 4;6(4):e01122. doi: 10.1128/mBio.01122-15. mBio. 2015. PMID: 26242627 Free PMC article.
-
Stress Beyond Translation: Poxviruses and More.Viruses. 2016 Jun 14;8(6):169. doi: 10.3390/v8060169. Viruses. 2016. PMID: 27314378 Free PMC article. Review.
-
Non-replicating Vaccinia Virus TianTan Strain (NTV) Translation Arrest of Viral Late Protein Synthesis Associated With Anti-viral Host Factor SAMD9.Front Cell Infect Microbiol. 2020 Mar 20;10:116. doi: 10.3389/fcimb.2020.00116. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 32266167 Free PMC article.
-
Innate immune recognition of poxviral vaccine vectors.Expert Rev Vaccines. 2011 Oct;10(10):1435-49. doi: 10.1586/erv.11.121. Expert Rev Vaccines. 2011. PMID: 21988308 Review.
Cited by
-
Novel biomarkers of resistance of pancreatic cancer cells to oncolytic vesicular stomatitis virus.Oncotarget. 2016 Sep 20;7(38):61601-61618. doi: 10.18632/oncotarget.11202. Oncotarget. 2016. PMID: 27533247 Free PMC article.
-
Rescue of a Vaccinia Virus Mutant Lacking IFN Resistance Genes K1L and C7L by the Parapoxvirus Orf Virus.Front Microbiol. 2020 Jul 28;11:1797. doi: 10.3389/fmicb.2020.01797. eCollection 2020. Front Microbiol. 2020. PMID: 32903701 Free PMC article.
-
High-throughput screening to enhance oncolytic virus immunotherapy.Oncolytic Virother. 2016 Apr 5;5:15-25. doi: 10.2147/OV.S66217. eCollection 2016. Oncolytic Virother. 2016. PMID: 27579293 Free PMC article. Review.
-
SAMD9L autoinflammatory or ataxia pancytopenia disease mutations activate cell-autonomous translational repression.Proc Natl Acad Sci U S A. 2021 Aug 24;118(34):e2110190118. doi: 10.1073/pnas.2110190118. Proc Natl Acad Sci U S A. 2021. PMID: 34417303 Free PMC article.
-
Emerging phenotypes linked to variants in SAMD9 and MIRAGE syndrome.Front Endocrinol (Lausanne). 2022 Aug 18;13:953707. doi: 10.3389/fendo.2022.953707. eCollection 2022. Front Endocrinol (Lausanne). 2022. PMID: 36060959 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
