

Cardiovascular defects in a mouse model of HOXA1 syndrome
- PMID: 21940751
- PMCID: PMC3235008
- DOI: 10.1093/hmg/ddr434
Cardiovascular defects in a mouse model of HOXA1 syndrome
Abstract
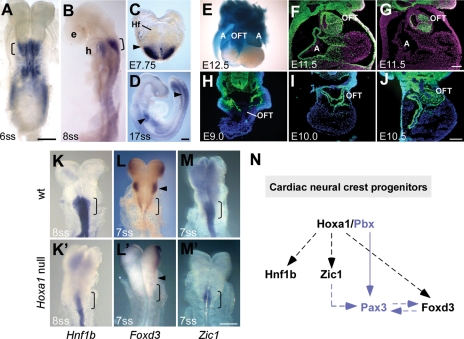
Congenital heart disease is one of the most common human birth defects, yet many genes and pathways regulating heart development remain unknown. A recent study in humans revealed that mutations in a single Hox gene, HOXA1 (Athabascan Brainstem Dysgenesis Syndrome, Bosley-Salih-Alorainy Syndrome), can cause severe cardiovascular malformations, some of which are lethal without surgical intervention. Since the discovery of the human syndromes, there have been no reports of any Hox mouse mutants with cardiac defects, hampering studies to explore the developmental causes of the human disease. In this study, we identify severe cardiovascular malformations in a Hox mouse model, which mimic the congenital heart defects in HOXA1 syndrome patients. Hoxa1 null mice show defects such as interrupted aortic arch, aberrant subclavian artery and Tetralogy of Fallot, demonstrating that Hoxa1 is required for patterning of the great arteries and outflow tract of the heart. We show that during early embryogenesis, Hoxa1 is expressed in precursors of cardiac neural crest cells (NCCs), which populate the heart. We further demonstrate that Hoxa1 acts upstream of several genes, important for neural crest specification. Thus, our data allow us to suggest a model in which Hoxa1 regulates heart development through its influence on cardiac NCCs, providing insight into the mechanisms underlying the human disease.
Figures
Similar articles
-
hoxa1a-Null Zebrafish as a Model for Studying HOXA1-Associated Heart Malformation in Bosley-Salih-Alorainy Syndrome.Biology (Basel). 2023 Jun 23;12(7):899. doi: 10.3390/biology12070899. Biology (Basel). 2023. PMID: 37508332 Free PMC article.
-
Hoxa1 and Hoxb1 are required for pharyngeal arch artery development.Mech Dev. 2017 Feb;143:1-8. doi: 10.1016/j.mod.2016.11.006. Epub 2016 Dec 10. Mech Dev. 2017. PMID: 27956219
-
Identification of novel Hoxa1 downstream targets regulating hindbrain, neural crest and inner ear development.Dev Biol. 2011 Sep 15;357(2):295-304. doi: 10.1016/j.ydbio.2011.06.042. Epub 2011 Jul 18. Dev Biol. 2011. PMID: 21784065 Free PMC article.
-
Role of the vascular endothelial growth factor isoforms in retinal angiogenesis and DiGeorge syndrome.Verh K Acad Geneeskd Belg. 2005;67(4):229-76. Verh K Acad Geneeskd Belg. 2005. PMID: 16334858 Review.
-
Molecular determinants of left and right outflow tract obstruction.Am J Med Genet. 2000 Winter;97(4):297-303. doi: 10.1002/1096-8628(200024)97:4<297::aid-ajmg1280>3.0.co;2-o. Am J Med Genet. 2000. PMID: 11376441 Review.
Cited by
-
Midbrain and hindbrain malformations: advances in clinical diagnosis, imaging, and genetics.Lancet Neurol. 2013 Apr;12(4):381-93. doi: 10.1016/S1474-4422(13)70024-3. Epub 2013 Mar 18. Lancet Neurol. 2013. PMID: 23518331 Free PMC article. Review.
-
Congenital cardiac anomalies and white matter injury.Trends Neurosci. 2015 Jun;38(6):353-63. doi: 10.1016/j.tins.2015.04.001. Epub 2015 May 1. Trends Neurosci. 2015. PMID: 25939892 Free PMC article. Review.
-
Single cell RNA-seq and ATAC-seq analysis of cardiac progenitor cell transition states and lineage settlement.Nat Commun. 2018 Nov 19;9(1):4877. doi: 10.1038/s41467-018-07307-6. Nat Commun. 2018. PMID: 30451828 Free PMC article.
-
hoxa1a-Null Zebrafish as a Model for Studying HOXA1-Associated Heart Malformation in Bosley-Salih-Alorainy Syndrome.Biology (Basel). 2023 Jun 23;12(7):899. doi: 10.3390/biology12070899. Biology (Basel). 2023. PMID: 37508332 Free PMC article.
-
Genetic and Developmental Contributors to Aortic Stenosis.Circ Res. 2021 Apr 30;128(9):1330-1343. doi: 10.1161/CIRCRESAHA.120.317978. Epub 2021 Apr 29. Circ Res. 2021. PMID: 33914609 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
