

The exopolygalacturonate lyase PelW and the oligogalacturonate lyase Ogl, two cytoplasmic enzymes of pectin catabolism in Erwinia chrysanthemi 3937
- PMID: 10383957
- PMCID: PMC93879
- DOI: 10.1128/JB.181.13.3912-3919.1999
The exopolygalacturonate lyase PelW and the oligogalacturonate lyase Ogl, two cytoplasmic enzymes of pectin catabolism in Erwinia chrysanthemi 3937
Abstract
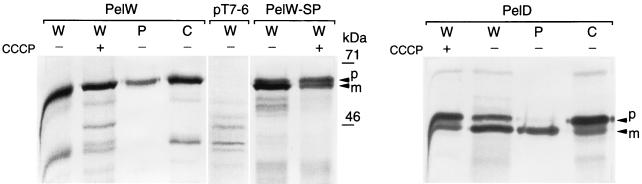
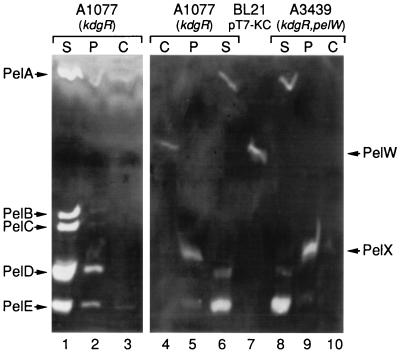
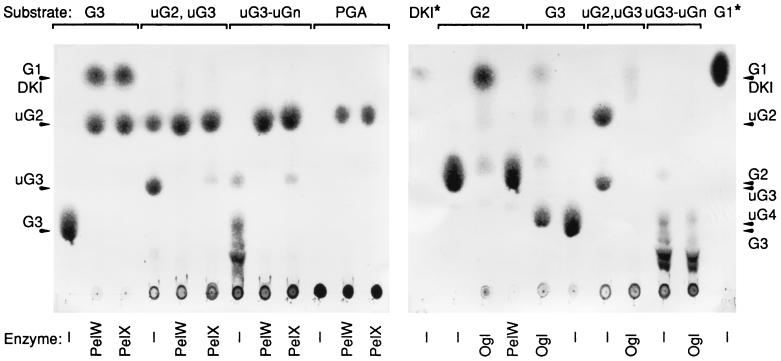
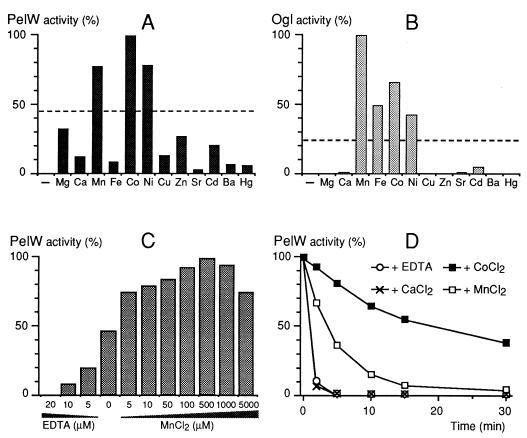
Erwinia chrysanthemi 3937 secretes into the external medium several pectinolytic enzymes, among which are eight isoenzymes of the endo-cleaving pectate lyases: PelA, PelB, PelC, PelD, and PelE (family 1); PelI (family 4); PelL (family 3); and PelZ (family 5). In addition, one exo-cleaving pectate lyase, PelX (family 3), has been found in the periplasm of E. chrysanthemi. The E. chrysanthemi 3937 gene kdgC has been shown to exhibit a high degree of similarity to the genes pelY of Yersinia pseudotuberculosis and pelB of Erwinia carotovora, which encode family 2 pectate lyases. However, no pectinolytic activity has been assigned to the KdgC protein. After verification of the corresponding nucleotide sequence, we cloned a longer DNA fragment and showed that this gene encodes a 553-amino-acid protein exhibiting an exo-cleaving pectate lyase activity. Thus, the kdgC gene was renamed pelW. PelW catalyzes the formation of unsaturated digalacturonates from polygalacturonate or short oligogalacturonates. PelW is located in the bacterial cytoplasm. In this compartment, PelW action could complete the degradation of pectic oligomers that was initiated by the extracellular or periplasmic pectinases and precede the action of the cytoplasmic oligogalacturonate lyase, Ogl. Both cytoplasmic pectinases, PelW and Ogl, seem to act in sequence during oligogalacturonate depolymerization, since oligomers longer than dimers are very poor substrates for Ogl but are good substrates for PelW. The estimated number of binding subsites for PelW is three, extending from subsite -2 to +1, while it is probably two for Ogl, extending from subsite -1 to +1. The activities of the two cytoplasmic lyases, PelW and Ogl, are dependent on the presence of divalent cations, since both enzymes are inhibited by EDTA. In contrast to the extracellular pectate lyases, Ca2+ is unable to restore the activity of PelW or Ogl, while several other cations, including Co2+, Mn2+, and Ni2+, can activate both cytoplasmic lyases.
Figures

Similar articles
-
Characterization of the exopolygalacturonate lyase PelX of Erwinia chrysanthemi 3937.J Bacteriol. 1999 Mar;181(5):1652-63. doi: 10.1128/JB.181.5.1652-1663.1999. J Bacteriol. 1999. PMID: 10049400 Free PMC article.
-
Characterization of the pelL gene encoding a novel pectate lyase of Erwinia chrysanthemi 3937.Mol Microbiol. 1995 Jun;16(6):1183-95. doi: 10.1111/j.1365-2958.1995.tb02341.x. Mol Microbiol. 1995. PMID: 8577252
-
Comparative analysis of the five major Erwinia chrysanthemi pectate lyases: enzyme characteristics and potential inhibitors.J Bacteriol. 1997 Apr;179(8):2503-11. doi: 10.1128/jb.179.8.2503-2511.1997. J Bacteriol. 1997. PMID: 9098045 Free PMC article.
-
Regulation of pectinolysis in Erwinia chrysanthemi.Annu Rev Microbiol. 1996;50:213-57. doi: 10.1146/annurev.micro.50.1.213. Annu Rev Microbiol. 1996. PMID: 8905080 Review.
-
Pectinolytic lyases: a comprehensive review of sources, category, property, structure, and catalytic mechanism of pectate lyases and pectin lyases.Bioresour Bioprocess. 2021 Aug 23;8(1):79. doi: 10.1186/s40643-021-00432-z. Bioresour Bioprocess. 2021. PMID: 38650254 Free PMC article. Review.
Cited by
-
Development of a Real-Time Pectic Oligosaccharide-Detecting Biosensor Using the Rapid and Flexible Computational Identification of Non-Disruptive Conjugation Sites (CINC) Biosensor Design Platform.Sensors (Basel). 2022 Jan 26;22(3):948. doi: 10.3390/s22030948. Sensors (Basel). 2022. PMID: 35161692 Free PMC article.
-
Biochemical Reconstruction of a Metabolic Pathway from a Marine Bacterium Reveals Its Mechanism of Pectin Depolymerization.Appl Environ Microbiol. 2018 Dec 13;85(1):e02114-18. doi: 10.1128/AEM.02114-18. Print 2019 Jan 1. Appl Environ Microbiol. 2018. PMID: 30341080 Free PMC article.
-
Comparison of expression, purification and characterization of a new pectate lyase from Phytophthora capsici using two different methods.BMC Biotechnol. 2011 Apr 6;11:32. doi: 10.1186/1472-6750-11-32. BMC Biotechnol. 2011. PMID: 21470403 Free PMC article.
-
Comparative Pathogenomic Analysis of Two Banana Pathogenic Dickeya Strains Isolated from China and the Philippines.Int J Mol Sci. 2022 Oct 22;23(21):12758. doi: 10.3390/ijms232112758. Int J Mol Sci. 2022. PMID: 36361548 Free PMC article.
-
KdgF, the missing link in the microbial metabolism of uronate sugars from pectin and alginate.Proc Natl Acad Sci U S A. 2016 May 31;113(22):6188-93. doi: 10.1073/pnas.1524214113. Epub 2016 May 16. Proc Natl Acad Sci U S A. 2016. PMID: 27185956 Free PMC article.
References
-
- Condemine G, Robert-Baudouy J. Analysis of an Erwinia chrysanthemi gene cluster involved in pectin degradation. Mol Microbiol. 1991;5:2191–2202. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
