

The Oxidative Stress-Induced Hypothetical Protein PG_0686 in Porphyromonas gingivalis W83 Is a Novel Diguanylate Cyclase
- PMID: 36719196
- PMCID: PMC10101095
- DOI: 10.1128/spectrum.04411-22
The Oxidative Stress-Induced Hypothetical Protein PG_0686 in Porphyromonas gingivalis W83 Is a Novel Diguanylate Cyclase
Abstract
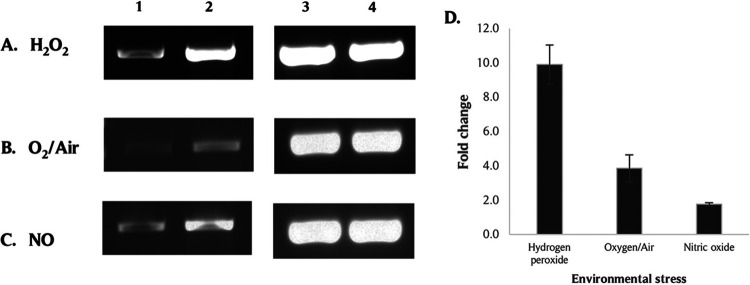
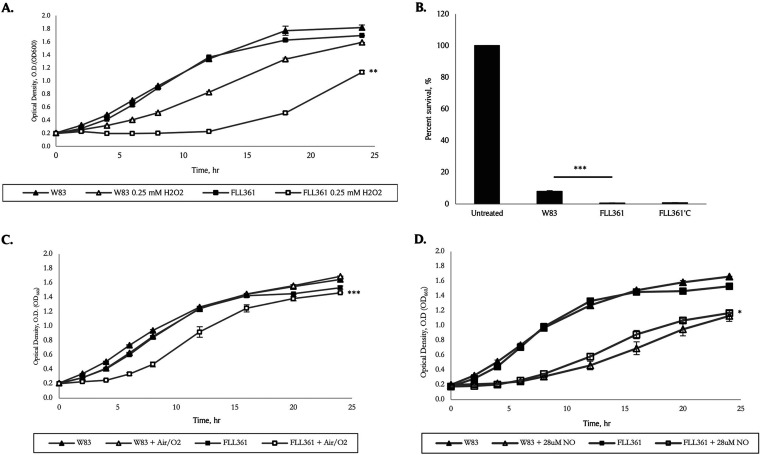
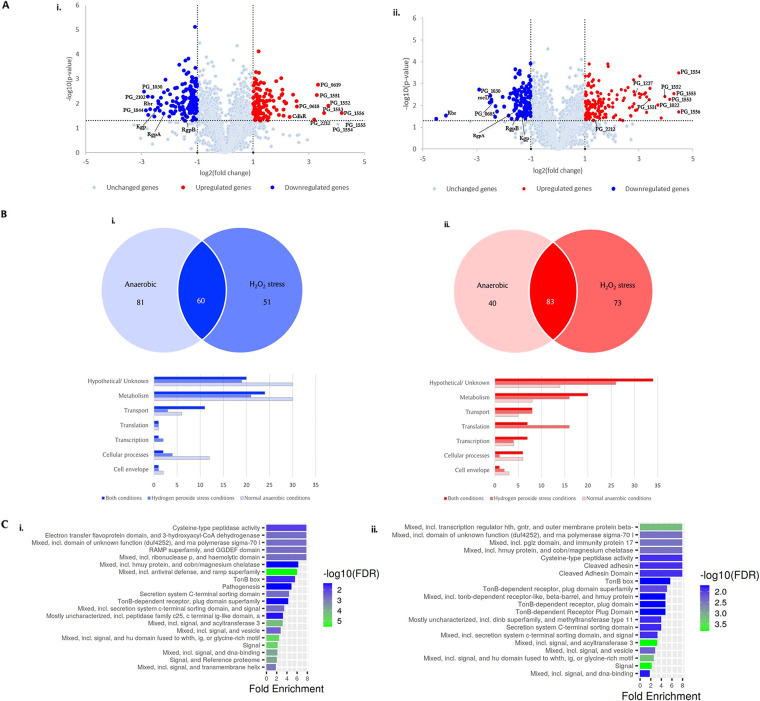
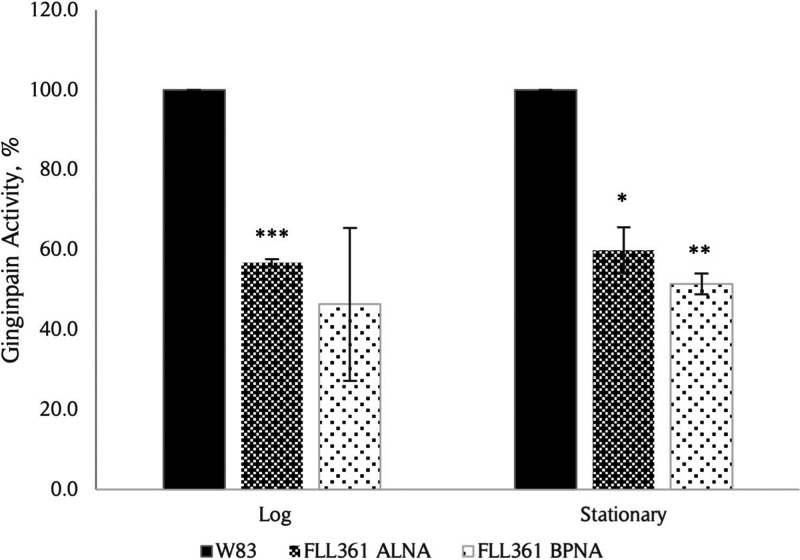
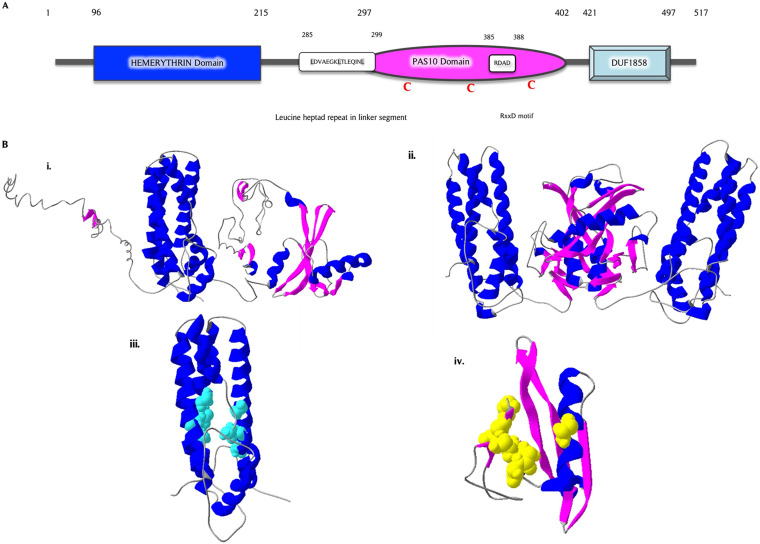
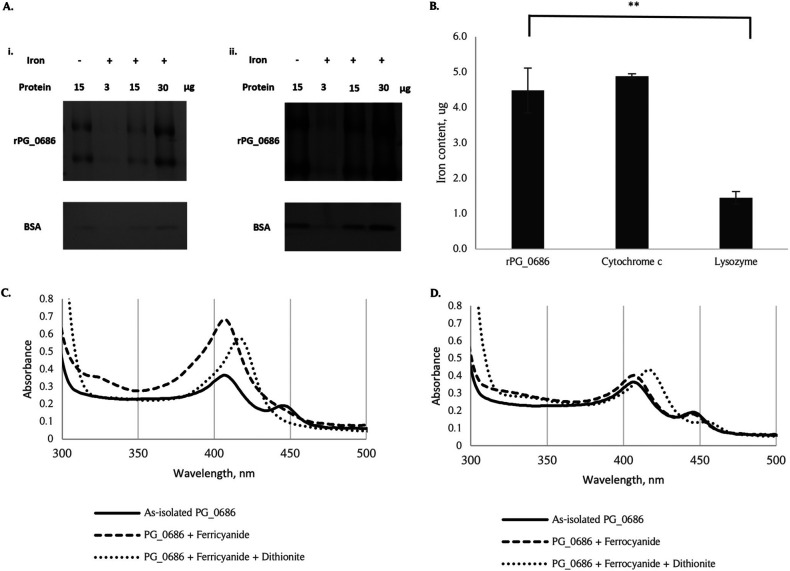
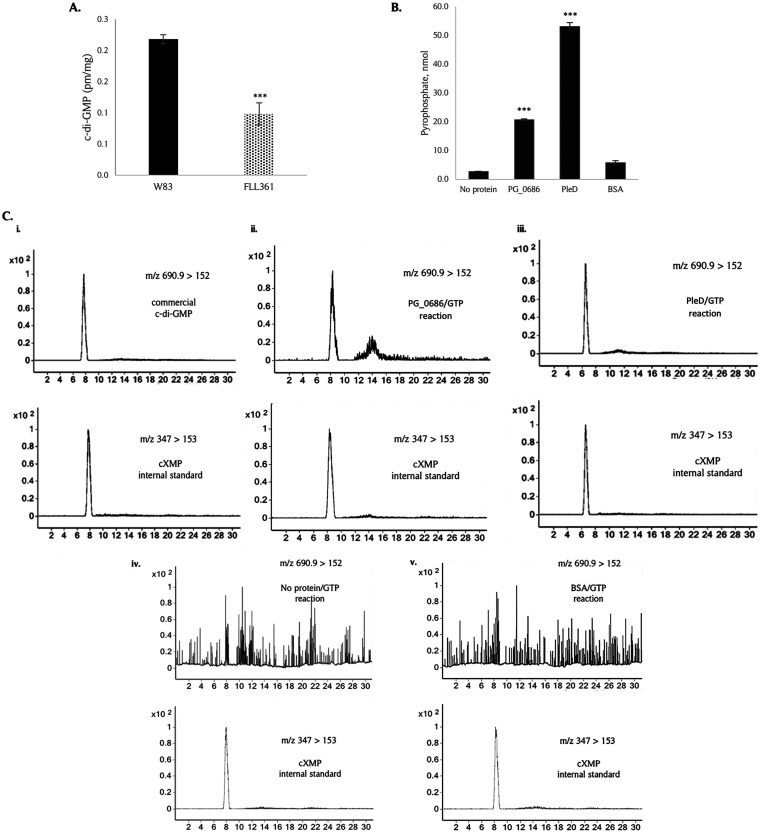
The survival/adaptation of Porphyromonas gingivalis to the inflammatory environment of the periodontal pocket requires an ability to overcome oxidative stress. Several functional classes of genes, depending on the severity and duration of the exposure, were induced in P. gingivalis under H2O2-induced oxidative stress. The PG_0686 gene was highly upregulated under prolonged oxidative stress. PG_0686, annotated as a hypothetical protein of unknown function, is a 60 kDa protein that carries several domains including hemerythrin, PAS10, and domain of unknown function (DUF)-1858. Although PG_0686 showed some relatedness to several diguanylate cyclases (DGCs), it is missing the classical conserved, active site sequence motif (GGD[/E]EF), commonly observed in other bacteria. PG_0686-related proteins are observed in other anaerobic bacterial species. The isogenic mutant P. gingivalis FLL361 (ΔPG_0686::ermF) showed increased sensitivity to H2O2, and decreased gingipain activity compared to the parental strain. Transcriptome analysis of P. gingivalis FLL361 showed the dysregulation of several gene clusters/operons, known oxidative stress resistance genes, and transcriptional regulators, including PG_2212, CdhR and PG_1181 that were upregulated under normal anaerobic conditions. The intracellular level of c-di-GMP in P. gingivalis FLL361 was significantly decreased compared to the parental strain. The purified recombinant PG_0686 (rPG_0686) protein catalyzed the formation of c-di-GMP from GTP. Collectively, our data suggest a global regulatory property for PG_0686 that may be part of an unconventional second messenger signaling system in P. gingivalis. Moreover, it may coordinately regulate a pathway(s) vital for protection against environmental stress, and is significant in the pathogenicity of P. gingivalis and other anaerobes. IMPORTANCE Porphyromonas gingivalis is an important etiological agent in periodontitis and other systemic diseases. There is still a gap in our understanding of the mechanisms that P. gingivalis uses to survive the inflammatory microenvironment of the periodontal pocket. The hypothetical PG_0686 gene was highly upregulated under prolonged oxidative stress. Although the tertiary structure of PG_0686 showed little relatedness to previously characterized diguanylate cyclases (DGCs), and does not contain the conserved GGD(/E)EF catalytic domain motif sequence, an ability to catalyze the formation of c-di-GMP from GTP is demonstrated. The second messenger pathway for c-di-GMP was previously predicted to be absent in P. gingivalis. PG_0686 paralogs are identified in other anaerobic bacteria. Thus, PG_0686 may represent a novel class of DGCs, which is yet to be characterized. In conclusion, we have shown, for the first time, evidence for the presence of c-di-GMP signaling with environmental stress protective function in P. gingivalis.
Keywords: PG_0686; Porphyromonas gingivalis; c-di GMP; diguanylate cyclase; oxidative stress.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Involvement of PG2212 zinc finger protein in the regulation of oxidative stress resistance in Porphyromonas gingivalis W83.J Bacteriol. 2014 Dec;196(23):4057-70. doi: 10.1128/JB.01907-14. Epub 2014 Sep 15. J Bacteriol. 2014. PMID: 25225267 Free PMC article.
-
Global Transcriptional Repression of Diguanylate Cyclases by MucR1 Is Essential for Sinorhizobium-Soybean Symbiosis.mBio. 2021 Oct 26;12(5):e0119221. doi: 10.1128/mBio.01192-21. Epub 2021 Oct 26. mBio. 2021. PMID: 34700374 Free PMC article.
-
Differential response of Porphyromonas gingivalis to varying levels and duration of hydrogen peroxide-induced oxidative stress.Microbiology (Reading). 2012 Oct;158(Pt 10):2465-2479. doi: 10.1099/mic.0.056416-0. Epub 2012 Jun 28. Microbiology (Reading). 2012. PMID: 22745271 Free PMC article.
-
Diguanylate Cyclases in Vibrio cholerae: Essential Regulators of Lifestyle Switching.Front Cell Infect Microbiol. 2020 Oct 22;10:582947. doi: 10.3389/fcimb.2020.582947. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 33194821 Free PMC article. Review.
-
Screening for Diguanylate Cyclase (DGC) Inhibitors Mitigating Bacterial Biofilm Formation.Front Chem. 2020 Apr 21;8:264. doi: 10.3389/fchem.2020.00264. eCollection 2020. Front Chem. 2020. PMID: 32373581 Free PMC article. Review.
Cited by
-
Friends and Foes in Alzheimer's Disease.Med Princ Pract. 2023;32(6):313-322. doi: 10.1159/000534400. Epub 2023 Oct 3. Med Princ Pract. 2023. PMID: 37788649 Free PMC article. Review.
-
Hemophore-like proteins of the HmuY family in the oral and gut microbiome: unraveling the mystery of their evolution.Microbiol Mol Biol Rev. 2024 Mar 27;88(1):e0013123. doi: 10.1128/mmbr.00131-23. Epub 2024 Feb 2. Microbiol Mol Biol Rev. 2024. PMID: 38305743 Free PMC article. Review.
-
Evolution of cyclic di-GMP signalling on a short and long term time scale.Microbiology (Reading). 2023 Jun;169(6):001354. doi: 10.1099/mic.0.001354. Microbiology (Reading). 2023. PMID: 37384391 Free PMC article.
References
-
- Wang X, Jia Y, Wen L, Mu W, Wu X, Liu T, Liu X, Fang J, Luan Y, Chen P, Gao J, Nguyen K-A, Cui J, Zeng G, Lan P, Chen Q, Cheng B, Wang Z. 2021. Porphyromonas gingivalis promotes colorectal carcinoma by activating the hematopoietic NLRP3 inflammasome. Cancer Res 81:2745–2759. doi:10.1158/0008-5472.CAN-20-3827. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
