

Bacteroides fragilis outer membrane vesicles preferentially activate innate immune receptors compared to their parent bacteria
- PMID: 36304461
- PMCID: PMC9592552
- DOI: 10.3389/fimmu.2022.970725
Bacteroides fragilis outer membrane vesicles preferentially activate innate immune receptors compared to their parent bacteria
Abstract
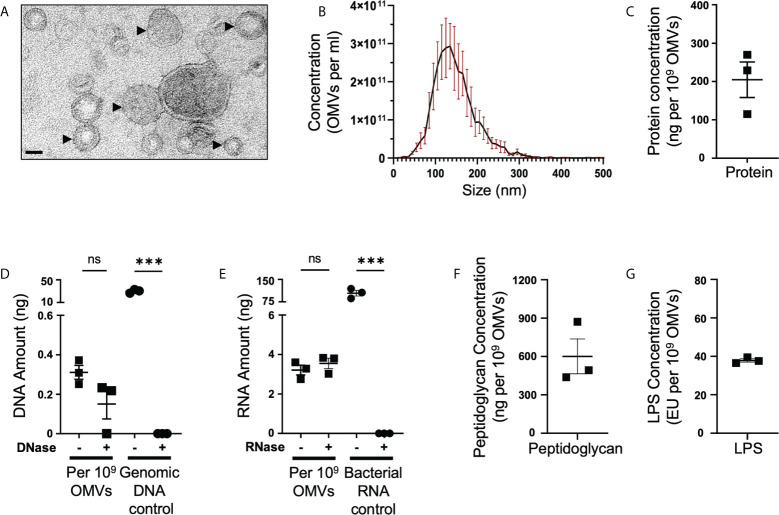
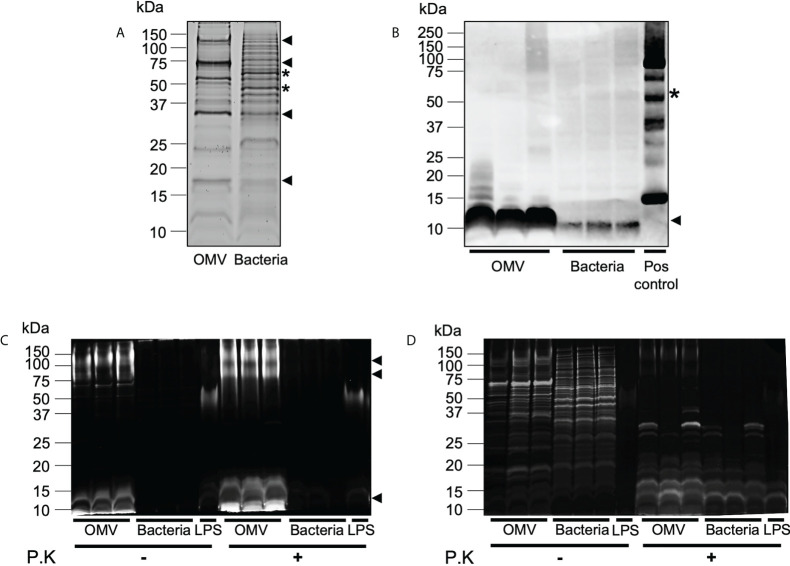
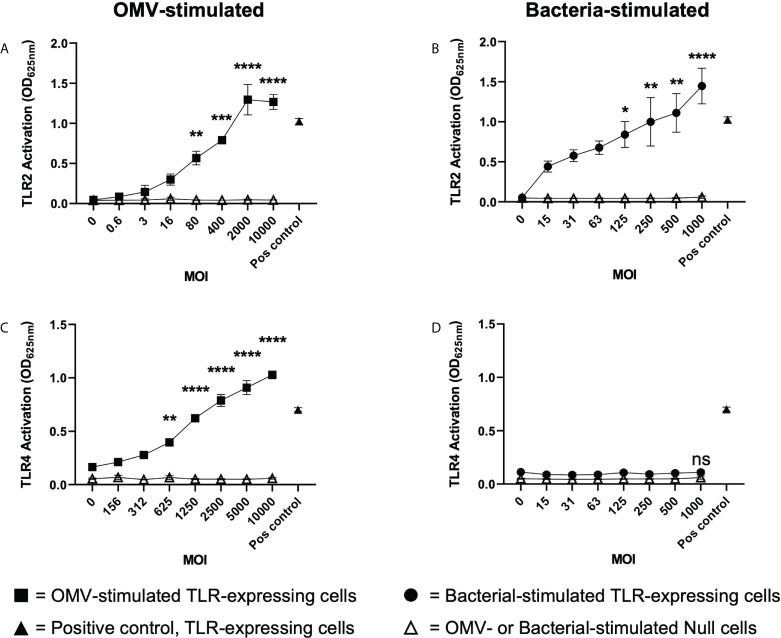
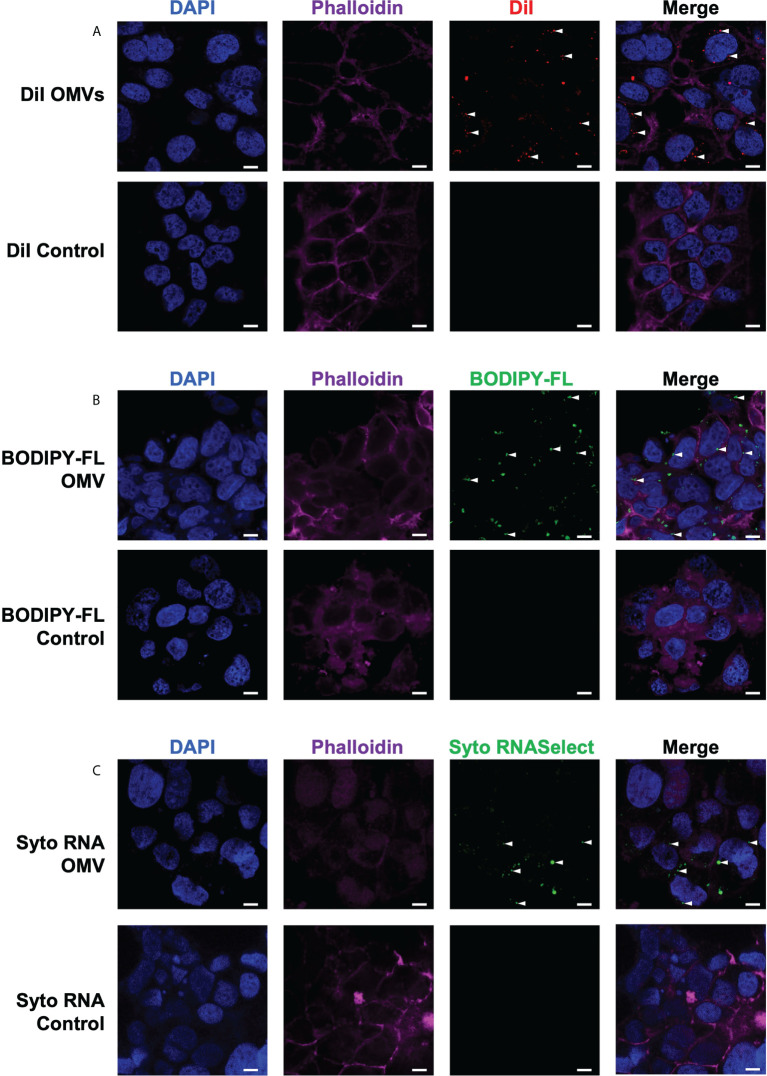
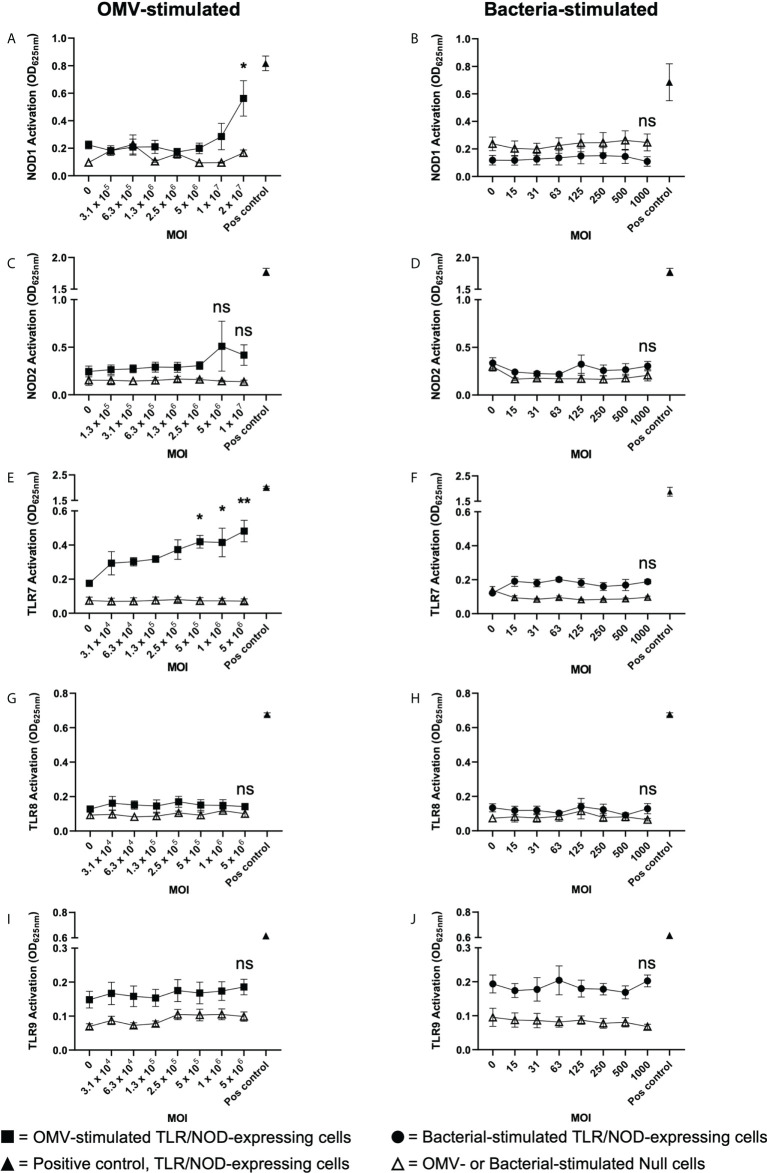
The release of bacterial membrane vesicles (BMVs) has become recognized as a key mechanism used by both pathogenic and commensal bacteria to activate innate immune responses in the host and mediate immunity. Outer membrane vesicles (OMVs) produced by Gram-negative bacteria can harbor various immunogenic cargo that includes proteins, nucleic acids and peptidoglycan, and the composition of OMVs strongly influences their ability to activate host innate immune receptors. Although various Gram-negative pathogens can produce OMVs that are enriched in immunogenic cargo compared to their parent bacteria, the ability of OMVs produced by commensal organisms to be enriched with immunostimulatory contents is only recently becoming known. In this study, we investigated the cargo associated with OMVs produced by the intestinal commensal Bacteroides fragilis and determined their ability to activate host innate immune receptors. Analysis of B. fragilis OMVs revealed that they packaged various biological cargo including proteins, DNA, RNA, lipopolysaccharides (LPS) and peptidoglycan, and that this cargo could be enriched in OMVs compared to their parent bacteria. We visualized the entry of B. fragilis OMVs into intestinal epithelial cells, in addition to the ability of B. fragilis OMVs to transport bacterial RNA and peptidoglycan cargo into Caco-2 epithelial cells. Using HEK-Blue reporter cell lines, we identified that B. fragilis OMVs could activate host Toll-like receptors (TLR)-2, TLR4, TLR7 and nucleotide-binding oligomerization domain-containing protein 1 (NOD1), whereas B. fragilis bacteria could only induce the activation of TLR2. Overall, our data demonstrates that B. fragilis OMVs activate a broader range of host innate immune receptors compared to their parent bacteria due to their enrichment of biological cargo and their ability to transport this cargo directly into host epithelial cells. These findings indicate that the secretion of OMVs by B. fragilis may facilitate immune crosstalk with host epithelial cells at the gastrointestinal surface and suggests that OMVs produced by commensal bacteria may preferentially activate host innate immune receptors at the mucosal gastrointestinal tract.
Keywords: Bacteroides fragilis; Commensals; NOD1; TLRs; bacterial membrane vesicles; epithelial cells; innate immunity; outer membrane vesicles (OMVs).
Copyright © 2022 Gilmore, Johnston, Bitto, Zavan, O'Brien-Simpson, Hill and Kaparakis-Liaskos.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Induction Effects of Bacteroides fragilis Derived Outer Membrane Vesicles on Toll Like Receptor 2, Toll Like Receptor 4 Genes Expression and Cytokines Concentration in Human Intestinal Epithelial Cells.Cell J. 2019 Apr;21(1):57-61. doi: 10.22074/cellj.2019.5750. Epub 2018 Nov 18. Cell J. 2019. PMID: 30507089 Free PMC article.
-
Detection of Bacterial Membrane Vesicles by NOD-Like Receptors.Int J Mol Sci. 2021 Jan 20;22(3):1005. doi: 10.3390/ijms22031005. Int J Mol Sci. 2021. PMID: 33498269 Free PMC article. Review.
-
Bacterial membrane vesicles deliver peptidoglycan to NOD1 in epithelial cells.Cell Microbiol. 2010 Mar;12(3):372-85. doi: 10.1111/j.1462-5822.2009.01404.x. Epub 2009 Nov 2. Cell Microbiol. 2010. PMID: 19888989
-
Staphylococcus aureus membrane vesicles contain immunostimulatory DNA, RNA and peptidoglycan that activate innate immune receptors and induce autophagy.J Extracell Vesicles. 2021 Apr;10(6):e12080. doi: 10.1002/jev2.12080. Epub 2021 Apr 1. J Extracell Vesicles. 2021. PMID: 33815695 Free PMC article.
-
Immunomodulatory roles and novel applications of bacterial membrane vesicles.Mol Immunol. 2021 Jun;134:72-85. doi: 10.1016/j.molimm.2021.02.027. Epub 2021 Mar 13. Mol Immunol. 2021. PMID: 33725501 Review.
Cited by
-
The role and mechanisms of gram-negative bacterial outer membrane vesicles in inflammatory diseases.Front Immunol. 2023 Jun 16;14:1157813. doi: 10.3389/fimmu.2023.1157813. eCollection 2023. Front Immunol. 2023. PMID: 37398647 Free PMC article. Review.
-
Characterization of the Inflammatory Response Evoked by Bacterial Membrane Vesicles in Intestinal Cells Reveals an RIPK2-Dependent Activation by Enterotoxigenic Escherichia coli Vesicles.Microbiol Spectr. 2023 Aug 17;11(4):e0111523. doi: 10.1128/spectrum.01115-23. Epub 2023 Jun 12. Microbiol Spectr. 2023. PMID: 37306596 Free PMC article.
-
Composition and functions of bacterial membrane vesicles.Nat Rev Microbiol. 2023 Jul;21(7):415-430. doi: 10.1038/s41579-023-00875-5. Epub 2023 Mar 17. Nat Rev Microbiol. 2023. PMID: 36932221 Review.
-
Gut commensals require Peyer's patches to induce protective systemic IgA responses.Res Sq [Preprint]. 2024 May 13:rs.3.rs-4220532. doi: 10.21203/rs.3.rs-4220532/v1. Res Sq. 2024. PMID: 38798510 Free PMC article. Preprint.
-
Gram-negative bacterial sRNAs encapsulated in OMVs: an emerging class of therapeutic targets in diseases.Front Cell Infect Microbiol. 2024 Jan 30;13:1305510. doi: 10.3389/fcimb.2023.1305510. eCollection 2023. Front Cell Infect Microbiol. 2024. PMID: 38983695 Free PMC article. Review.
References
-
- Kadurugamuwa JL, Beveridge TJ. Virulence factors are released from Pseudomonas aeruginosa in association with membrane vesicles during normal growth and exposure to gentamicin: a novel mechanism of enzyme secretion. J Bacteriol (1995) 177(14):3998–4008. doi: 10.1128/jb.177.14.3998-4008.1995 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous