

Molecular Basis for the Interaction Between AP4 β4 and its Accessory Protein, Tepsin
- PMID: 26756312
- PMCID: PMC4805503
- DOI: 10.1111/tra.12375
Molecular Basis for the Interaction Between AP4 β4 and its Accessory Protein, Tepsin
Abstract
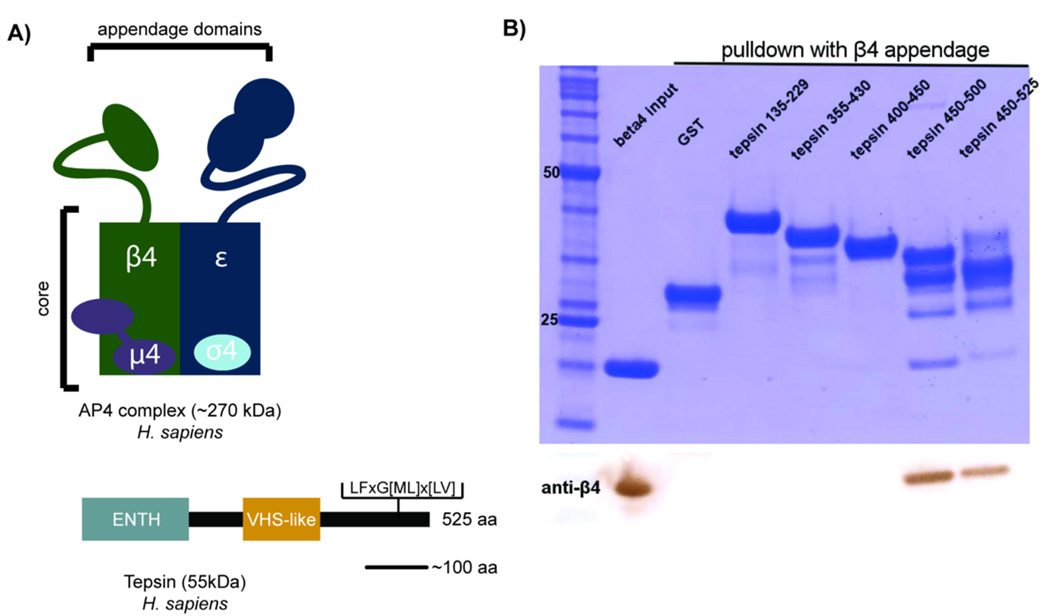
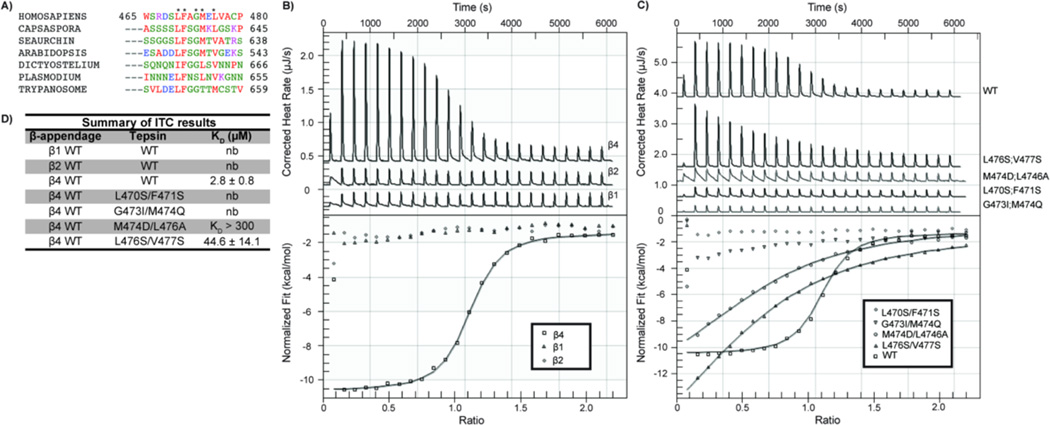
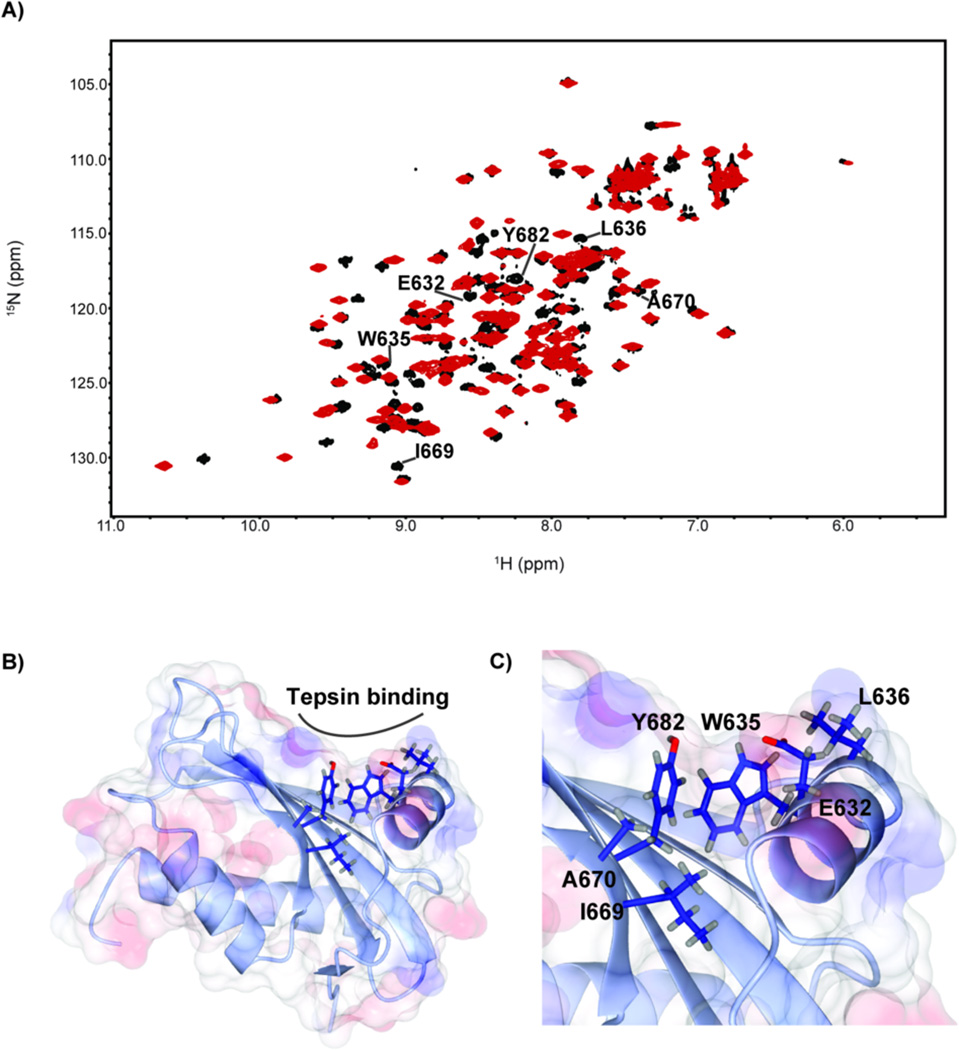
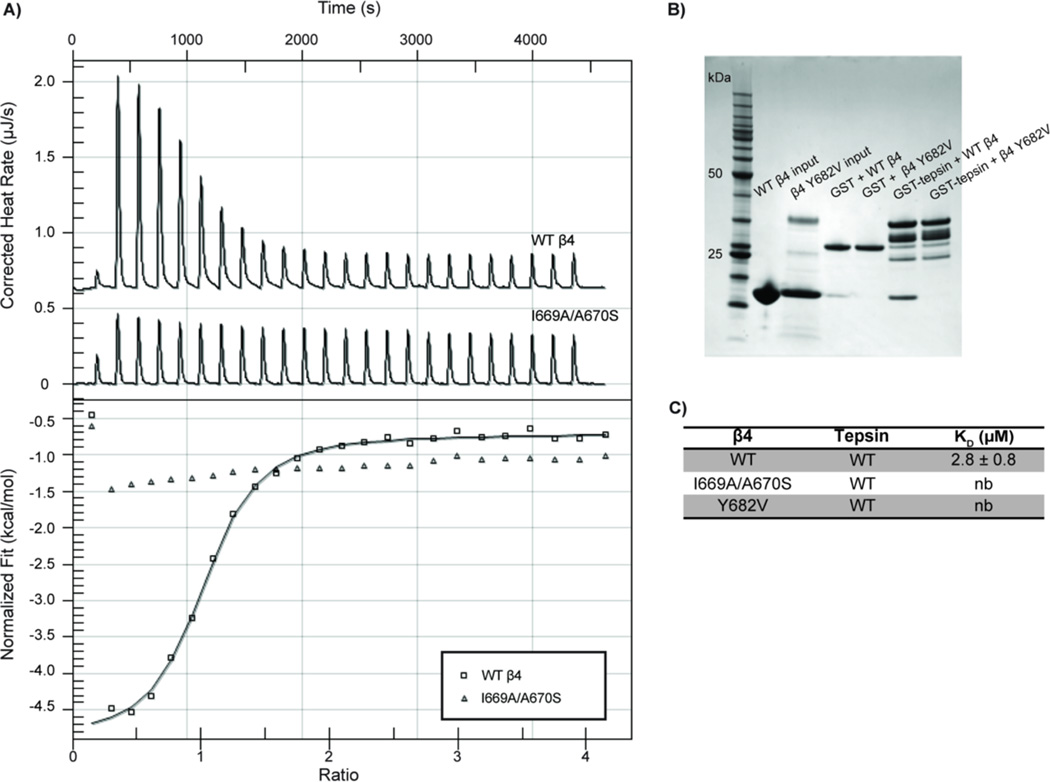
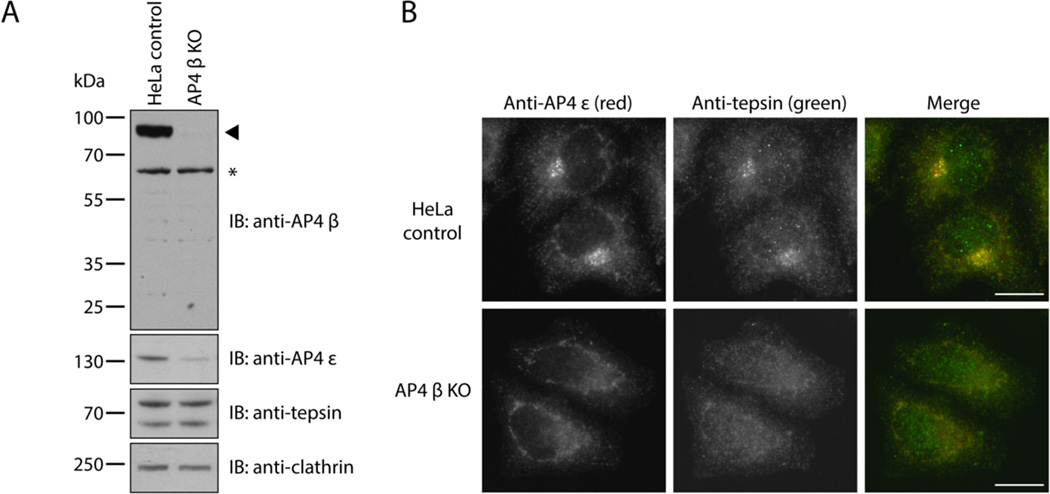
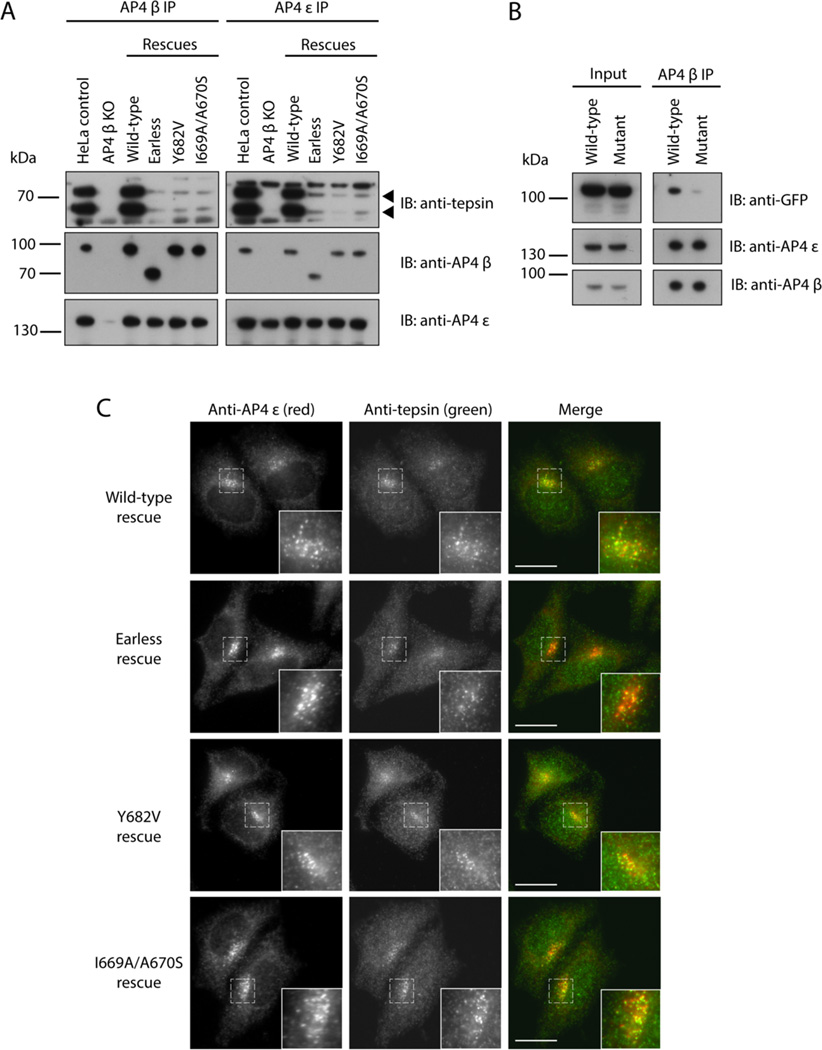
The adaptor protein 4 (AP4) complex (ϵ/β4/μ4/σ4 subunits) forms a non-clathrin coat on vesicles departing the trans-Golgi network. AP4 biology remains poorly understood, in stark contrast to the wealth of molecular data available for the related clathrin adaptors AP1 and AP2. AP4 is important for human health because mutations in any AP4 subunit cause severe neurological problems, including intellectual disability and progressive spastic para- or tetraplegias. We have used a range of structural, biochemical and biophysical approaches to determine the molecular basis for how the AP4 β4 C-terminal appendage domain interacts with tepsin, the only known AP4 accessory protein. We show that tepsin harbors a hydrophobic sequence, LFxG[M/L]x[L/V], in its unstructured C-terminus, which binds directly and specifically to the C-terminal β4 appendage domain. Using nuclear magnetic resonance chemical shift mapping, we define the binding site on the β4 appendage by identifying residues on the surface whose signals are perturbed upon titration with tepsin. Point mutations in either the tepsin LFxG[M/L]x[L/V] sequence or in its cognate binding site on β4 abolish in vitro binding. In cells, the same point mutations greatly reduce the amount of tepsin that interacts with AP4. However, they do not abolish the binding between tepsin and AP4 completely, suggesting the existence of additional interaction sites between AP4 and tepsin. These data provide one of the first detailed mechanistic glimpses at AP4 coat assembly and should provide an entry point for probing the role of AP4-coated vesicles in cell biology, and especially in neuronal function.
Keywords: adaptor protein complexes; biochemistry; cell biology; membrane trafficking; non-clathrin coats; structural biology; vesicle coats.
© 2016 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
Structure and evolution of ENTH and VHS/ENTH-like domains in tepsin.Traffic. 2017 Sep;18(9):590-603. doi: 10.1111/tra.12499. Traffic. 2017. PMID: 28691777 Free PMC article.
-
Bivalent Motif-Ear Interactions Mediate the Association of the Accessory Protein Tepsin with the AP-4 Adaptor Complex.J Biol Chem. 2015 Dec 25;290(52):30736-49. doi: 10.1074/jbc.M115.683409. Epub 2015 Nov 5. J Biol Chem. 2015. PMID: 26542808 Free PMC article.
-
Structural and functional characterization of cargo-binding sites on the μ4-subunit of adaptor protein complex 4.PLoS One. 2014 Feb 3;9(2):e88147. doi: 10.1371/journal.pone.0088147. eCollection 2014. PLoS One. 2014. PMID: 24498434 Free PMC article.
-
The role of AP-4 in cargo export from the trans-Golgi network and hereditary spastic paraplegia.Biochem Soc Trans. 2020 Oct 30;48(5):1877-1888. doi: 10.1042/BST20190664. Biochem Soc Trans. 2020. PMID: 33084855 Review.
-
Conformational regulation of AP1 and AP2 clathrin adaptor complexes.Traffic. 2019 Oct;20(10):741-751. doi: 10.1111/tra.12677. Epub 2019 Aug 6. Traffic. 2019. PMID: 31313456 Free PMC article. Review.
Cited by
-
The adaptor protein chaperone AAGAB stabilizes AP-4 complex subunits.Mol Biol Cell. 2022 Oct 1;33(12):ar109. doi: 10.1091/mbc.E22-05-0177. Epub 2022 Aug 17. Mol Biol Cell. 2022. PMID: 35976721 Free PMC article.
-
Structure and evolution of ENTH and VHS/ENTH-like domains in tepsin.Traffic. 2017 Sep;18(9):590-603. doi: 10.1111/tra.12499. Traffic. 2017. PMID: 28691777 Free PMC article.
-
Cargo Sorting at the trans-Golgi Network for Shunting into Specific Transport Routes: Role of Arf Small G Proteins and Adaptor Complexes.Cells. 2019 Jun 3;8(6):531. doi: 10.3390/cells8060531. Cells. 2019. PMID: 31163688 Free PMC article. Review.
-
Pre-clinical development of AP4B1 gene replacement therapy for hereditary spastic paraplegia type 47.EMBO Mol Med. 2024 Nov;16(11):2882-2917. doi: 10.1038/s44321-024-00148-5. Epub 2024 Oct 2. EMBO Mol Med. 2024. PMID: 39358605 Free PMC article.
-
Evolutionary origins and specialisation of membrane transport.Curr Opin Cell Biol. 2018 Aug;53:70-76. doi: 10.1016/j.ceb.2018.06.001. Epub 2018 Jun 19. Curr Opin Cell Biol. 2018. PMID: 29929066 Free PMC article. Review.
References
-
- Owen DJ, Evans PR. A structural explanation for the recognition of tyrosine-based endocytotic signals. Science (80-) [Internet] 1998;282:1327–1332. Available from: http://www.ncbi.nlm.nih.gov/pubmed/9812899. - PMC - PubMed
-
- Owen DJ, Vallis Y, Noble MEM, Hunter JB, Dafforn TR, Evans PR, McMahon HT. A Structural Explanation for the Binding of Multiple Ligands by the α-adaptin Appendage Domain. Cell. 1999;97:805–815. - PubMed
-
- Traub LM, Downs MA, Westrich JL, Fremont DH. Crystal structure of the alpha appendage of AP-2 reveals a recruitment platform for clathrin-coat assembly. Proc Natl Acad Sci U S A [Internet] 1999;96:8907–8912. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=17706&tool=pmc.... - PMC - PubMed
-
- Owen DJ, Vallis Y, Pearse BM, McMahon HT, Evans PR. The structure and function of the β2-adaptin appendage domain. EMBO J [Internet] 2000;19:4216–4227. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=302036&tool=pm.... - PMC - PubMed
-
- Collins BM, McCoy AJ, Kent HM, Evans PR, Owen DJ. Molecular architecture and functional model of the endocytic AP2 complex. Cell [Internet] 2002;109:523–535. Available from: http://www.ncbi.nlm.nih.gov/pubmed/12086608. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
