

Missense dopamine transporter mutations associate with adult parkinsonism and ADHD
- PMID: 24911152
- PMCID: PMC4071392
- DOI: 10.1172/JCI73778
Missense dopamine transporter mutations associate with adult parkinsonism and ADHD
Abstract
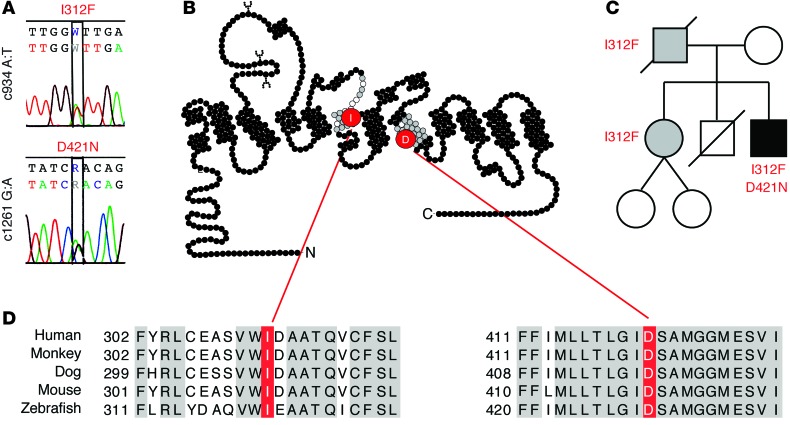
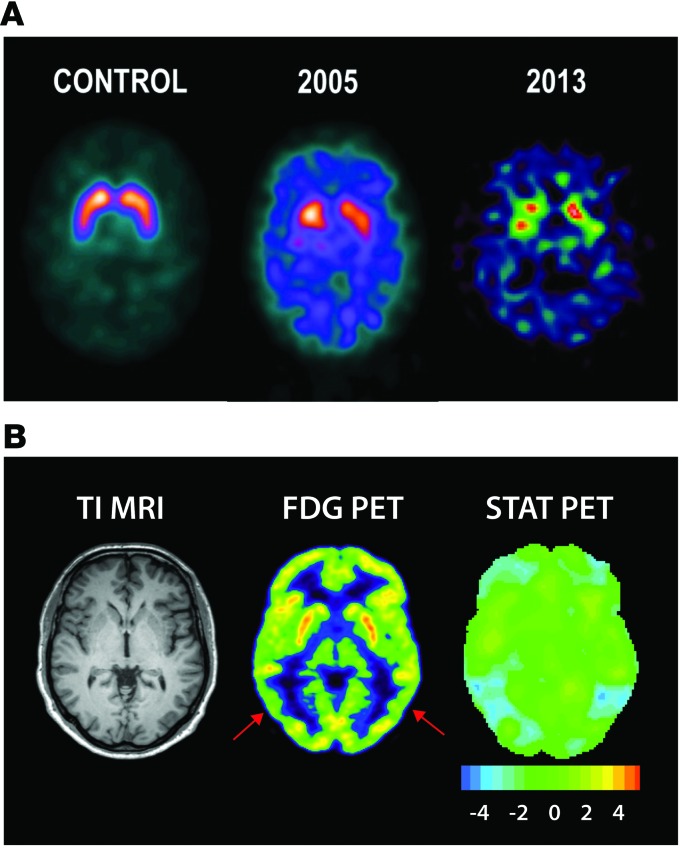
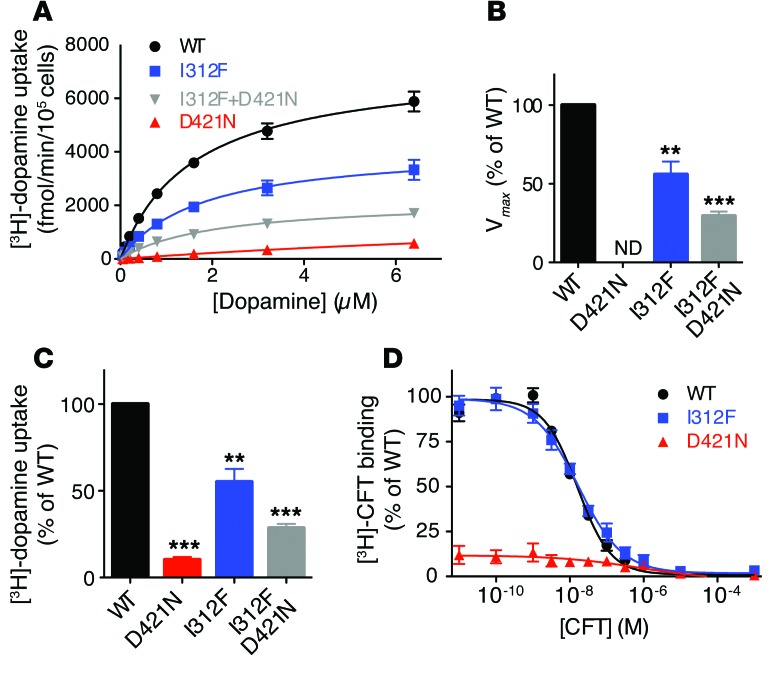
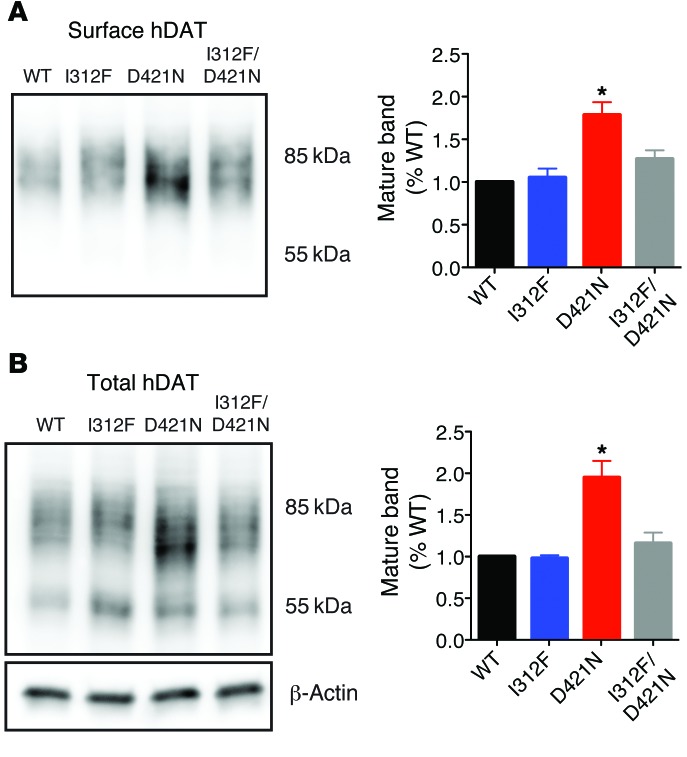
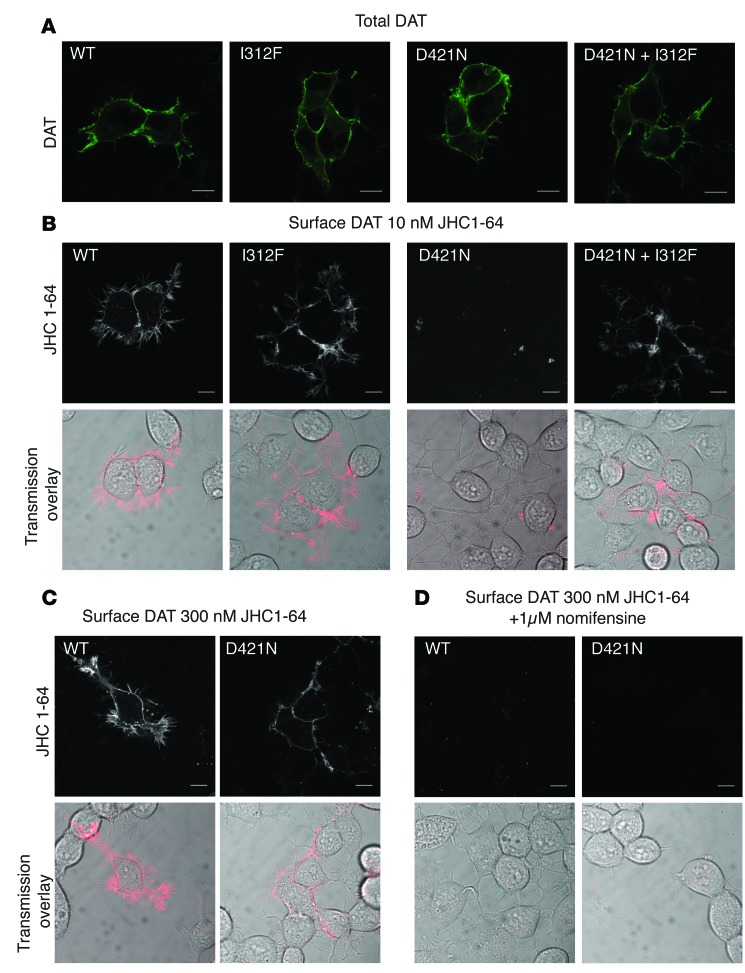
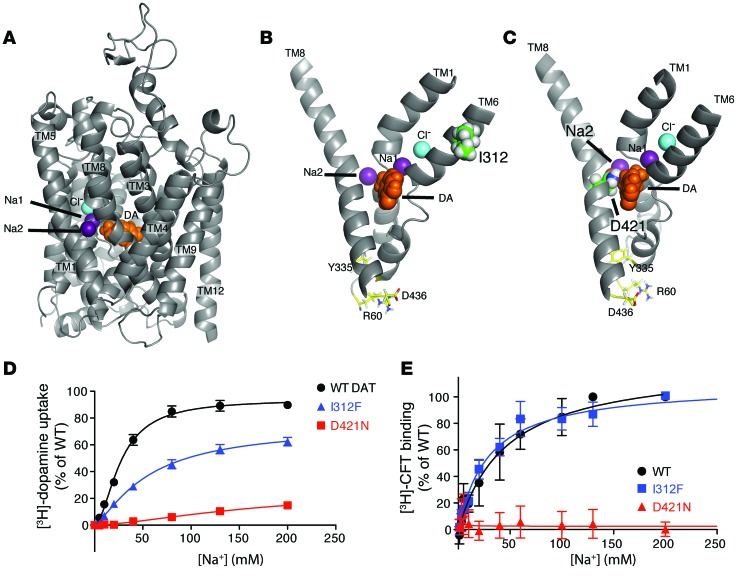
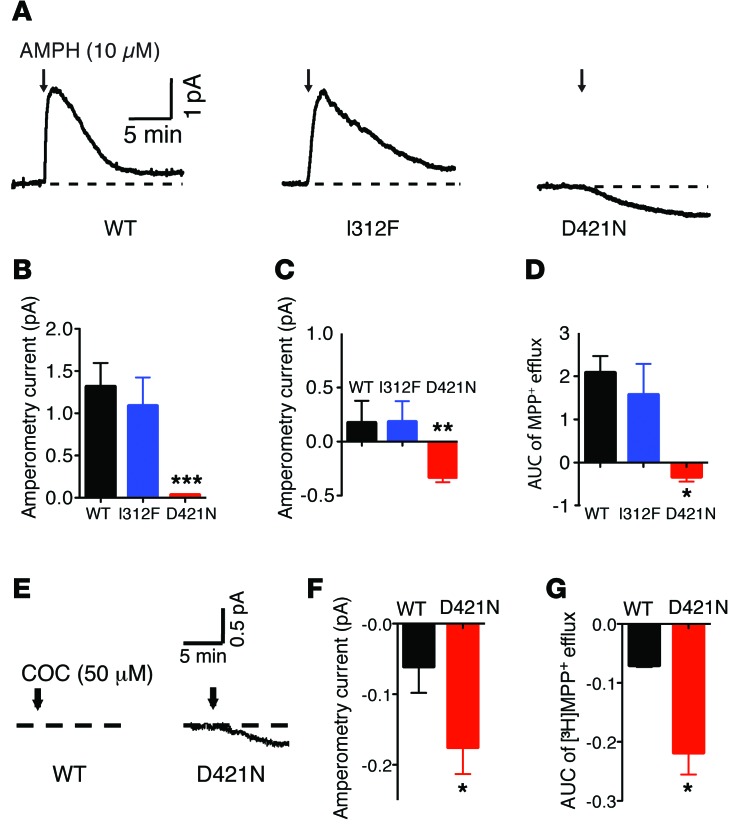
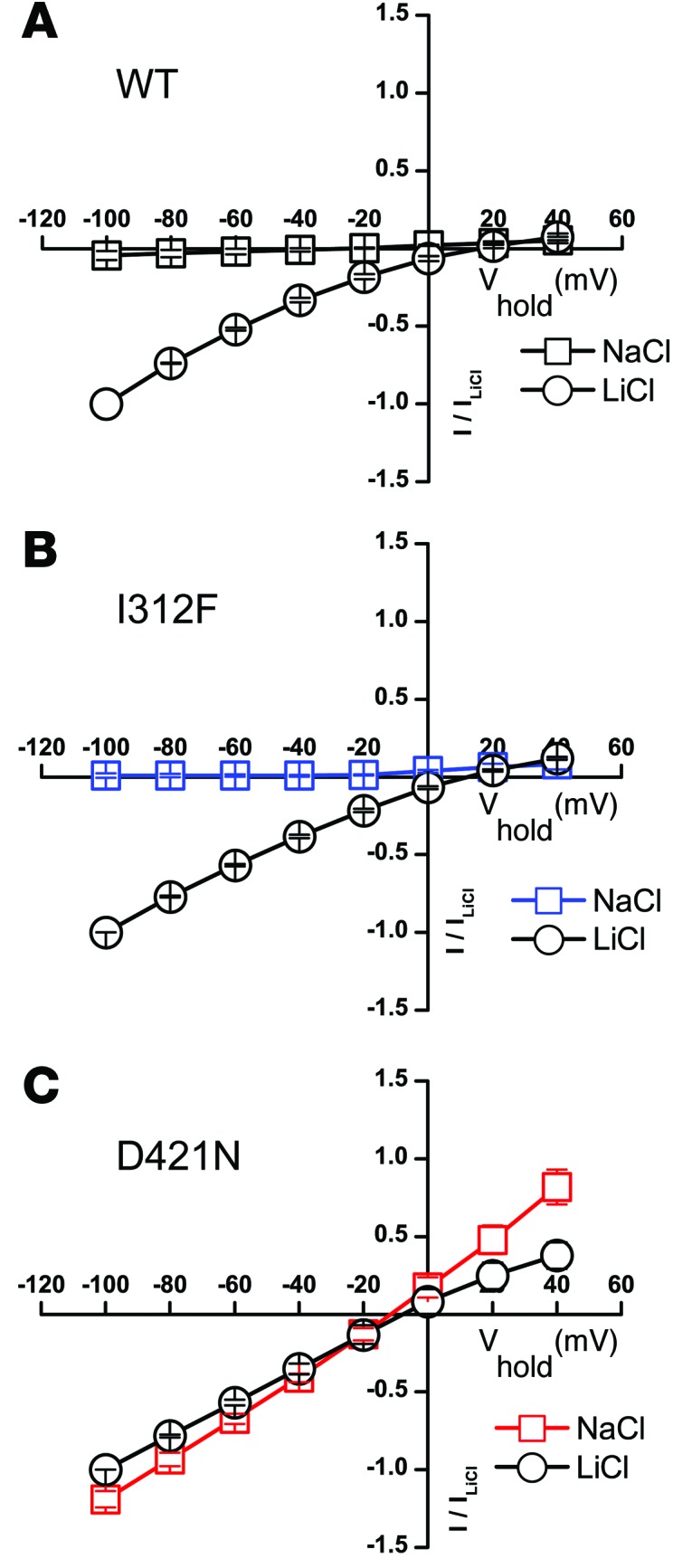
Parkinsonism and attention deficit hyperactivity disorder (ADHD) are widespread brain disorders that involve disturbances of dopaminergic signaling. The sodium-coupled dopamine transporter (DAT) controls dopamine homeostasis, but its contribution to disease remains poorly understood. Here, we analyzed a cohort of patients with atypical movement disorder and identified 2 DAT coding variants, DAT-Ile312Phe and a presumed de novo mutant DAT-Asp421Asn, in an adult male with early-onset parkinsonism and ADHD. According to DAT single-photon emission computed tomography (DAT-SPECT) scans and a fluoro-deoxy-glucose-PET/MRI (FDG-PET/MRI) scan, the patient suffered from progressive dopaminergic neurodegeneration. In heterologous cells, both DAT variants exhibited markedly reduced dopamine uptake capacity but preserved membrane targeting, consistent with impaired catalytic activity. Computational simulations and uptake experiments suggested that the disrupted function of the DAT-Asp421Asn mutant is the result of compromised sodium binding, in agreement with Asp421 coordinating sodium at the second sodium site. For DAT-Asp421Asn, substrate efflux experiments revealed a constitutive, anomalous efflux of dopamine, and electrophysiological analyses identified a large cation leak that might further perturb dopaminergic neurotransmission. Our results link specific DAT missense mutations to neurodegenerative early-onset parkinsonism. Moreover, the neuropsychiatric comorbidity provides additional support for the idea that DAT missense mutations are an ADHD risk factor and suggests that complex DAT genotype and phenotype correlations contribute to different dopaminergic pathologies.
Figures
Similar articles
-
Identifying dominant-negative actions of a dopamine transporter variant in patients with parkinsonism and neuropsychiatric disease.JCI Insight. 2021 Sep 22;6(18):e151496. doi: 10.1172/jci.insight.151496. JCI Insight. 2021. PMID: 34375312 Free PMC article.
-
Neuropsychiatric disease-associated genetic variants of the dopamine transporter display heterogeneous molecular phenotypes.J Biol Chem. 2018 May 11;293(19):7250-7262. doi: 10.1074/jbc.RA118.001753. Epub 2018 Mar 20. J Biol Chem. 2018. PMID: 29559554 Free PMC article.
-
Homozygous loss-of-function mutations in the gene encoding the dopamine transporter are associated with infantile parkinsonism-dystonia.J Clin Invest. 2009 Jun;119(6):1595-603. doi: 10.1172/JCI39060. Epub 2009 May 26. J Clin Invest. 2009. PMID: 19478460 Free PMC article.
-
SPECT and PET of the dopamine transporter in attention-deficit/hyperactivity disorder.Expert Rev Neurother. 2008 Apr;8(4):611-25. doi: 10.1586/14737175.8.4.611. Expert Rev Neurother. 2008. PMID: 18416663 Review.
-
ADHD and the dopamine transporter: are there reasons to pay attention?Handb Exp Pharmacol. 2006;(175):373-415. doi: 10.1007/3-540-29784-7_17. Handb Exp Pharmacol. 2006. PMID: 16722244 Review.
Cited by
-
Alpha-synuclein modulates dopamine neurotransmission.J Chem Neuroanat. 2017 Oct;83-84:41-49. doi: 10.1016/j.jchemneu.2016.06.001. Epub 2016 Jun 19. J Chem Neuroanat. 2017. PMID: 27334403 Free PMC article. Review.
-
Gene therapy for neurotransmitter-related disorders.J Inherit Metab Dis. 2024 Jan;47(1):176-191. doi: 10.1002/jimd.12697. J Inherit Metab Dis. 2024. PMID: 38221762 Free PMC article. Review.
-
Membrane transporters as mediators of synaptic dopamine dynamics: implications for disease.Eur J Neurosci. 2017 Jan;45(1):20-33. doi: 10.1111/ejn.13357. Epub 2016 Sep 2. Eur J Neurosci. 2017. PMID: 27520881 Free PMC article. Review.
-
Heterozygote Dopamine Transporter Knockout Rats Display Enhanced Cocaine Locomotion in Adolescent Females.Int J Mol Sci. 2022 Dec 6;23(23):15414. doi: 10.3390/ijms232315414. Int J Mol Sci. 2022. PMID: 36499749 Free PMC article.
-
Disturbed neurotransmitter homeostasis in ether lipid deficiency.Hum Mol Genet. 2019 Jun 15;28(12):2046-2061. doi: 10.1093/hmg/ddz040. Hum Mol Genet. 2019. PMID: 30759250 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
