

Band 3 Courcouronnes (Ser667Phe): a trafficking mutant differentially rescued by wild-type band 3 and glycophorin A
- PMID: 18174378
- PMCID: PMC2605348
- DOI: 10.1182/blood-2007-07-099473
Band 3 Courcouronnes (Ser667Phe): a trafficking mutant differentially rescued by wild-type band 3 and glycophorin A
Abstract
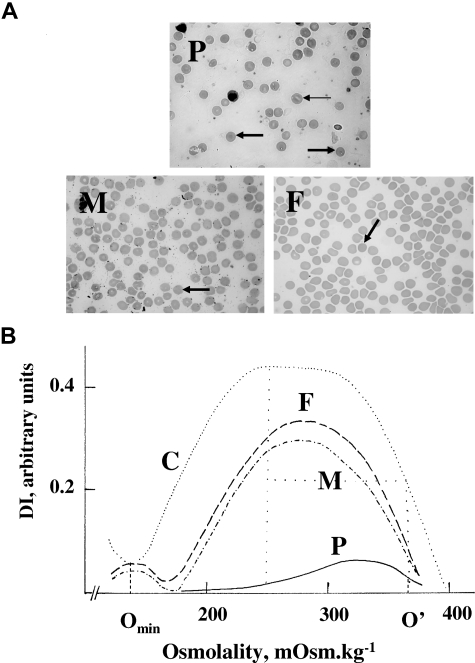
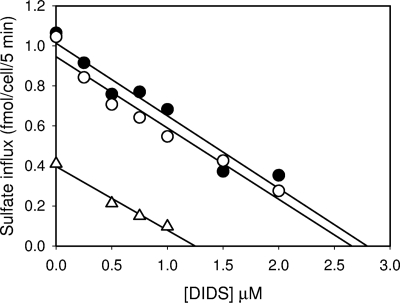
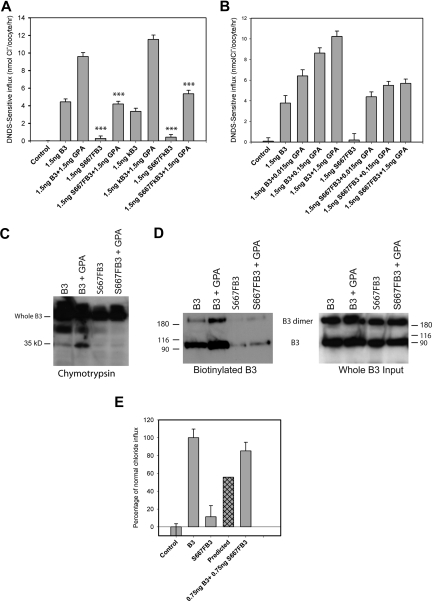
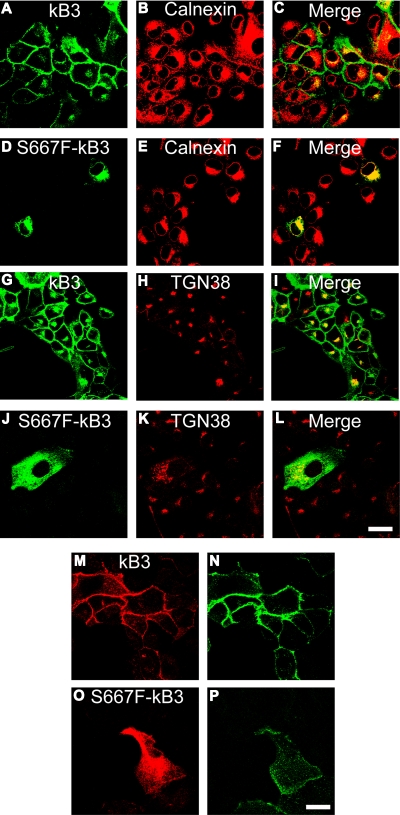
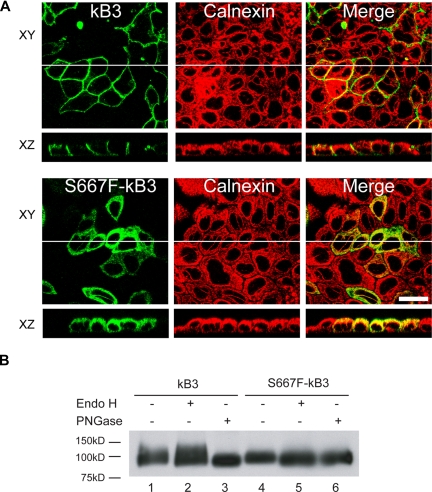
We describe a mutation in human erythrocyte band 3 (anion exchanger 1; SLC4A1) causing both hereditary spherocytosis and distal renal tubular acidosis. The proband developed a transfusion-dependent, hemolytic anemia following birth. Immunoblotting showed band 3 was reduced to approximately 35% of wildtype; other proteins of the band 3/Rh macrocomplex were also reduced. DNA sequence analysis revealed a novel homozygous mutation, c.2000C>T, leading to the amino acid substitution Ser667Phe. The parents were heterozygous for the same mutation. Sulfate influx in the patient's erythrocytes was approximately 40% wild type. The mutant band 3 produced very little chloride influx when expressed in Xenopus oocytes. Influx was partially rescued by coexpression of glycophorin A and also rescued by coexpression of wild-type band 3. At 2 years of age, an ammonium chloride challenge showed the child has incomplete distal renal tubular acidosis (dRTA). Stable expression of mutant kidney band 3 in both nonpolarized and polarized Madin-Darby canine kidney cells showed that most of the mutant protein was retained in the endoplasmic reticulum. Overall our results suggest that the Ser667Phe does not affect the anion transport function of band 3, but causes a trafficking defect in both erythrocytes and kidney cells.
Figures

Similar articles
-
Band 3 Edmonton I, a novel mutant of the anion exchanger 1 causing spherocytosis and distal renal tubular acidosis.Biochem J. 2010 Feb 24;426(3):379-88. doi: 10.1042/BJ20091525. Biochem J. 2010. PMID: 20028337
-
Novel AE1 mutations in recessive distal renal tubular acidosis. Loss-of-function is rescued by glycophorin A.J Clin Invest. 1998 Dec 15;102(12):2173-9. doi: 10.1172/JCI4836. J Clin Invest. 1998. PMID: 9854053 Free PMC article.
-
A novel missense mutation in AE1 causing autosomal dominant distal renal tubular acidosis retains normal transport function but is mistargeted in polarized epithelial cells.J Biol Chem. 2004 Apr 2;279(14):13833-8. doi: 10.1074/jbc.M400188200. Epub 2004 Jan 20. J Biol Chem. 2004. PMID: 14734552
-
The association between familial distal renal tubular acidosis and mutations in the red cell anion exchanger (band 3, AE1) gene.Biochem Cell Biol. 1998;76(5):723-8. doi: 10.1139/bcb-76-5-723. Biochem Cell Biol. 1998. PMID: 10353704 Review.
-
Band 3, the human red cell chloride/bicarbonate anion exchanger (AE1, SLC4A1), in a structural context.Biochim Biophys Acta. 2016 Jul;1858(7 Pt A):1507-32. doi: 10.1016/j.bbamem.2016.03.030. Epub 2016 Apr 6. Biochim Biophys Acta. 2016. PMID: 27058983 Review.
Cited by
-
The sorting of blood group active proteins during enucleation.ISBT Sci Ser. 2015 Apr 1;10(Suppl 1):163-168. doi: 10.1111/voxs.12127. ISBT Sci Ser. 2015. PMID: 26640516 Free PMC article.
-
Molecular physiology and genetics of Na+-independent SLC4 anion exchangers.J Exp Biol. 2009 Jun;212(Pt 11):1672-83. doi: 10.1242/jeb.029454. J Exp Biol. 2009. PMID: 19448077 Free PMC article. Review.
-
Reticulocyte Maturation and Variant Red Blood Cells.Front Physiol. 2022 Mar 7;13:834463. doi: 10.3389/fphys.2022.834463. eCollection 2022. Front Physiol. 2022. PMID: 35356079 Free PMC article.
-
A Ser725Arg mutation in Band 3 abolishes transport function and leads to anemia and renal tubular acidosis.Blood. 2018 Apr 12;131(15):1759-1763. doi: 10.1182/blood-2018-01-827725. Epub 2018 Feb 26. Blood. 2018. PMID: 29483102 Free PMC article. No abstract available.
-
Cell physiology and molecular mechanism of anion transport by erythrocyte band 3/AE1.Am J Physiol Cell Physiol. 2021 Dec 1;321(6):C1028-C1059. doi: 10.1152/ajpcell.00275.2021. Epub 2021 Oct 20. Am J Physiol Cell Physiol. 2021. PMID: 34669510 Free PMC article. Review.
References
-
- Tanner MJA. Band 3 anion exchanger and its involvement in erythrocyte and kidney disorders. Curr Opin Hematol. 2002;9:133–139. - PubMed
-
- Gallagher PG, Forget BG. Hematologically important mutations: band 3 and protein 4. 2 variants in hereditary spherocytosis. Blood Cells Mol Dis. 1997;23:417–421. - PubMed
-
- Wrong O, Bruce LJ, Unwin RJ, Toye AM, Tanner MJ. Band 3 mutations, distal renal tubular acidosis, and Southeast Asian ovalocytosis. Kidney Int. 2002;62:10–19. - PubMed
-
- Toye AM. Defective kidney anion-exchanger 1 (AE1, Band 3) trafficking in dominant distal renal tubular acidosis (dRTA). Biochem Soc Symp. 2005;72:47–63. - PubMed
-
- Peters LL, Shivdasani RA, Liu SC, et al. Anion exchanger 1 (Band 3) is required to prevent erythrocyte surface loss but not to form the membrane skeleton. Cell. 1996;86:917–927. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
