

Novel loci and biomedical consequences of iron homoeostasis variation
- PMID: 39643614
- PMCID: PMC11624196
- DOI: 10.1038/s42003-024-07115-3
Novel loci and biomedical consequences of iron homoeostasis variation
Abstract
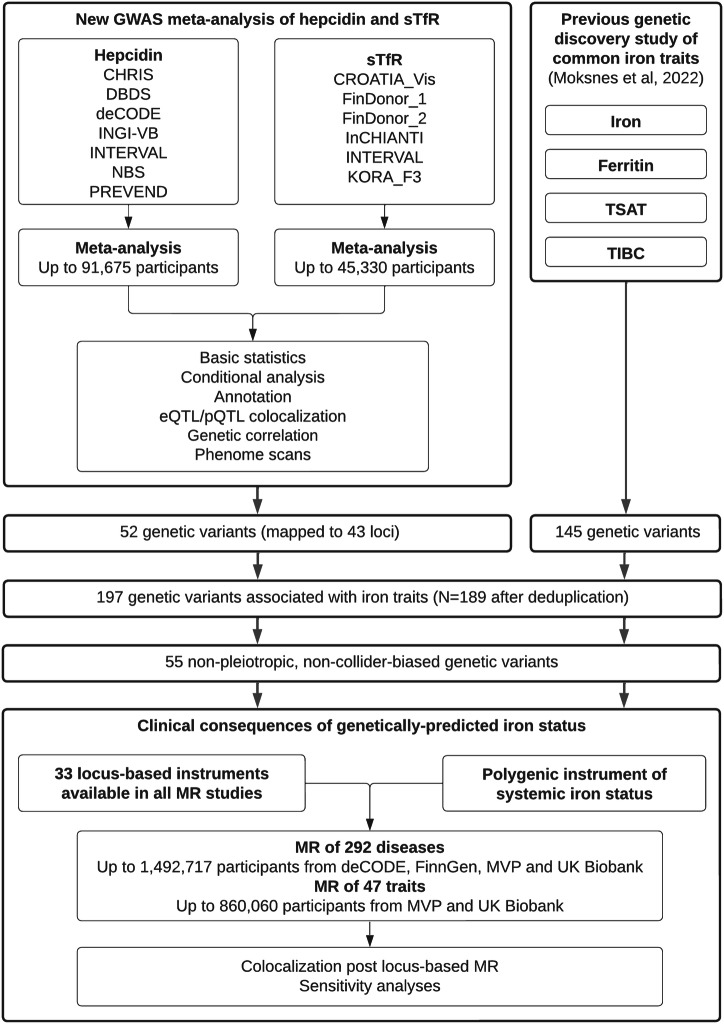
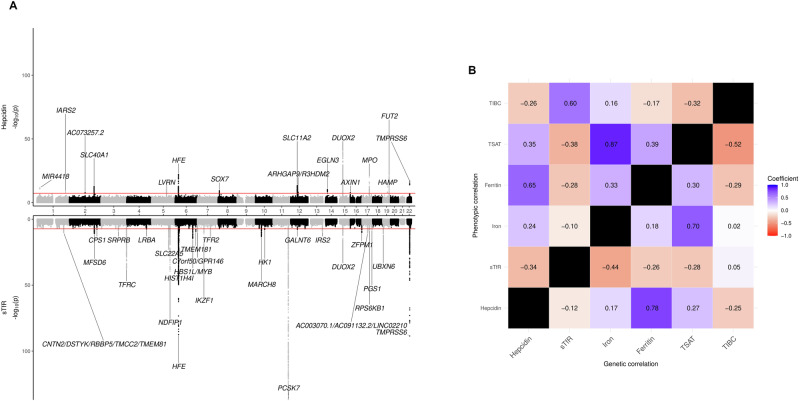
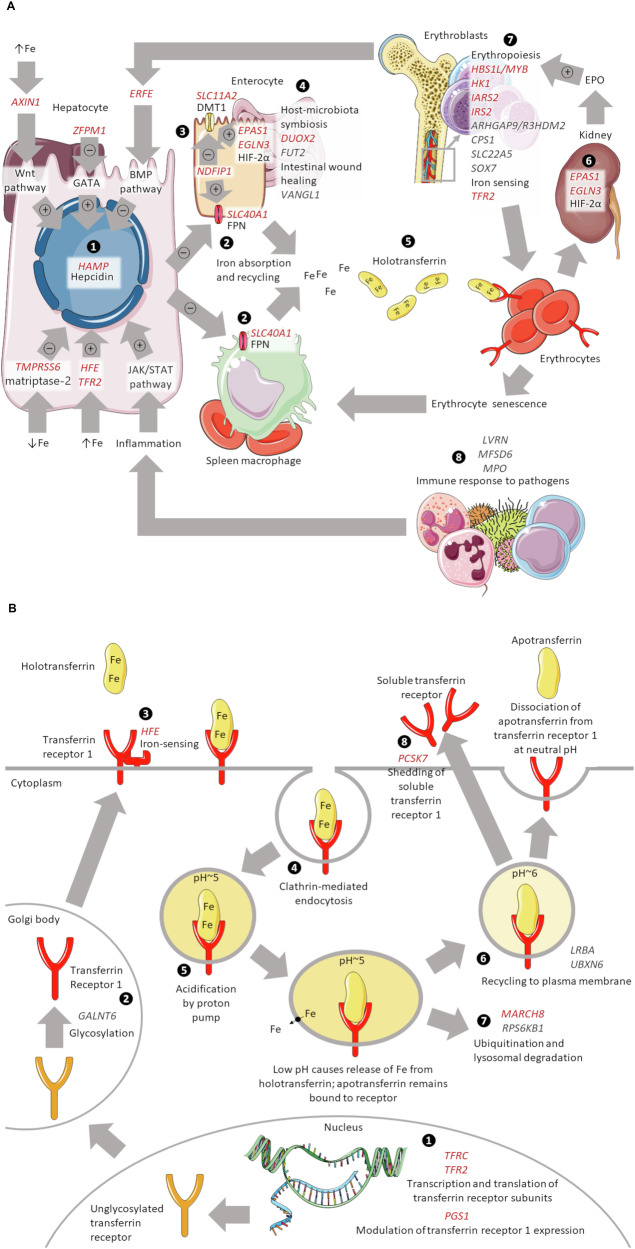
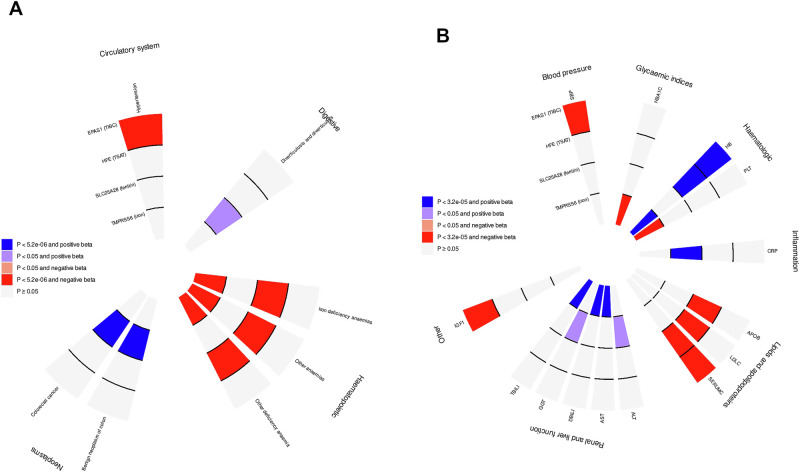
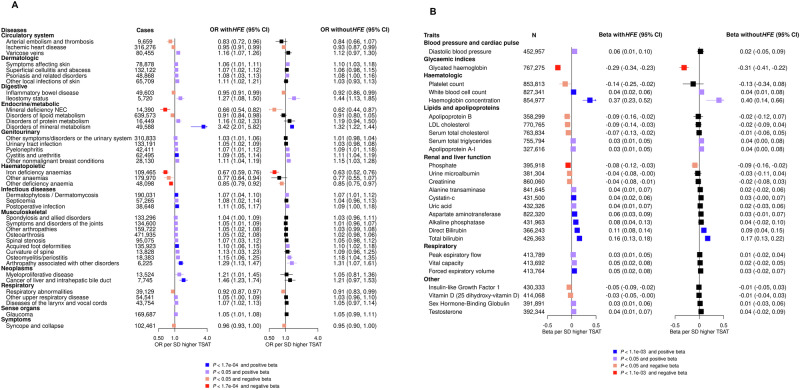
Iron homoeostasis is tightly regulated, with hepcidin and soluble transferrin receptor (sTfR) playing significant roles. However, the genetic determinants of these traits and the biomedical consequences of iron homoeostasis variation are unclear. In a meta-analysis of 12 cohorts involving 91,675 participants, we found 43 genomic loci associated with either hepcidin or sTfR concentration, of which 15 previously unreported. Mapping to putative genes indicated involvement in iron-trait expression, erythropoiesis, immune response and cellular trafficking. Mendelian randomisation of 292 disease outcomes in 1,492,717 participants revealed associations of iron-related loci and iron status with selected health outcomes across multiple domains. These associations were largely driven by HFE, which was associated with the largest iron variation. Our findings enhance understanding of iron homoeostasis and its biomedical consequences, suggesting that lifelong exposure to higher iron levels is likely associated with lower risk of anaemia-related disorders and higher risk of genitourinary, musculoskeletal, infectious and neoplastic diseases.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: R.S. is currently employed at Astra Zeneca. N.V. is an employee and stockholder of Regeneron Pharmaceuticals. J.Da. serves on scientific advisory boards for AstraZeneca, Novartis, and UK Biobank, and has received multiple grants from academic, charitable, and industry sources outside of the submitted work. All other authors declare no competing interests.
Figures
Similar articles
-
Hepcidin expression is associated with increased γ-secretase-mediated cleavage of neogenin in the liver.J Biol Chem. 2024 Dec;300(12):107927. doi: 10.1016/j.jbc.2024.107927. Epub 2024 Oct 24. J Biol Chem. 2024. PMID: 39454953 Free PMC article.
-
A phenome-wide association and Mendelian randomisation study of alcohol use variants in a diverse cohort comprising over 3 million individuals.EBioMedicine. 2024 May;103:105086. doi: 10.1016/j.ebiom.2024.105086. Epub 2024 Apr 4. EBioMedicine. 2024. PMID: 38580523 Free PMC article.
-
Cross-Ancestry Investigation of Venous Thromboembolism Genomic Predictors.Circulation. 2022 Oct 18;146(16):1225-1242. doi: 10.1161/CIRCULATIONAHA.122.059675. Epub 2022 Sep 26. Circulation. 2022. PMID: 36154123 Free PMC article.
-
Strategies to improve smoking cessation rates in primary care.Cochrane Database Syst Rev. 2021 Sep 6;9(9):CD011556. doi: 10.1002/14651858.CD011556.pub2. Cochrane Database Syst Rev. 2021. PMID: 34693994 Free PMC article. Review.
-
Depressing time: Waiting, melancholia, and the psychoanalytic practice of care.In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. PMID: 36137063 Free Books & Documents. Review.
References
-
- Camaschella, C. Iron-deficiency anemia. N. Engl. J. Med.372, 1832–1843 (2015). - PubMed
-
- Hentze, M. W., Muckenthaler, M. U., Galy, B. & Camaschella, C. Two to tango: regulation of Mammalian iron metabolism. Cell142, 24–38 (2010). - PubMed
-
- Beguin, Y. Soluble transferrin receptor for the evaluation of erythropoiesis and iron status. Clin. Chim. Acta329, 9–22 (2003). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
