

Killer prey: Ecology reverses bacterial predation
- PMID: 38261596
- PMCID: PMC10805292
- DOI: 10.1371/journal.pbio.3002454
Killer prey: Ecology reverses bacterial predation
Abstract
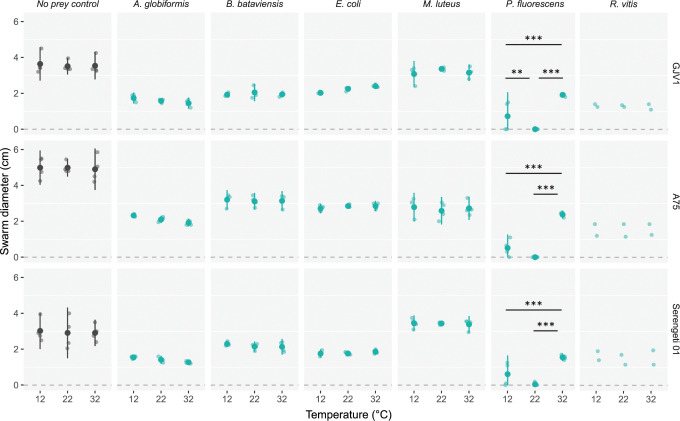
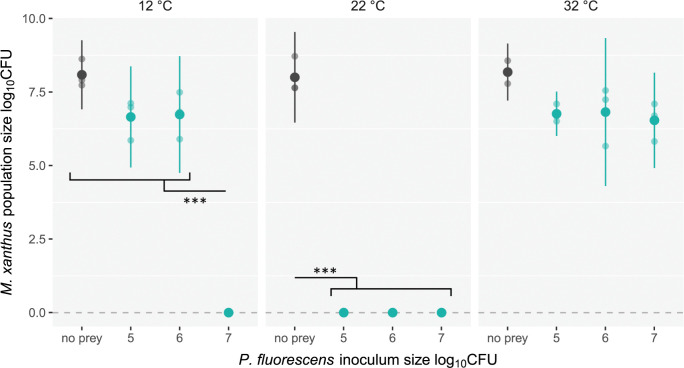
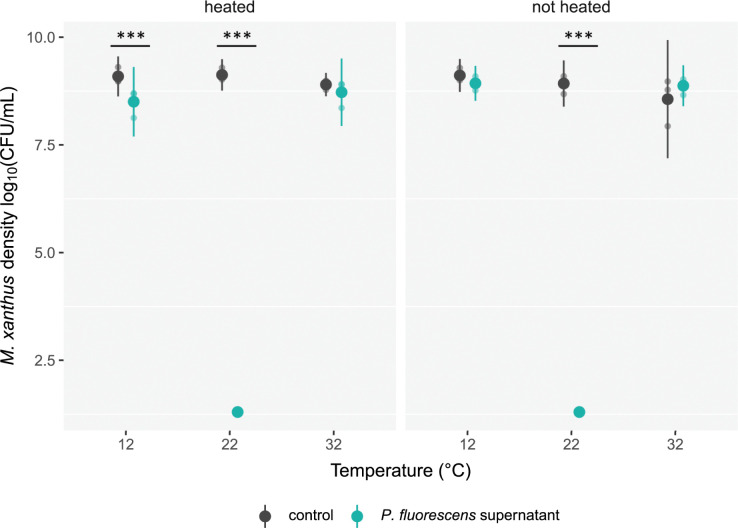
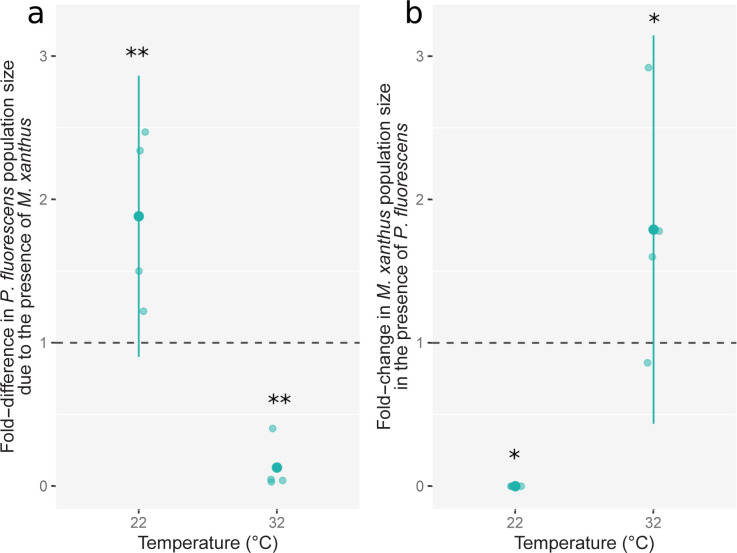
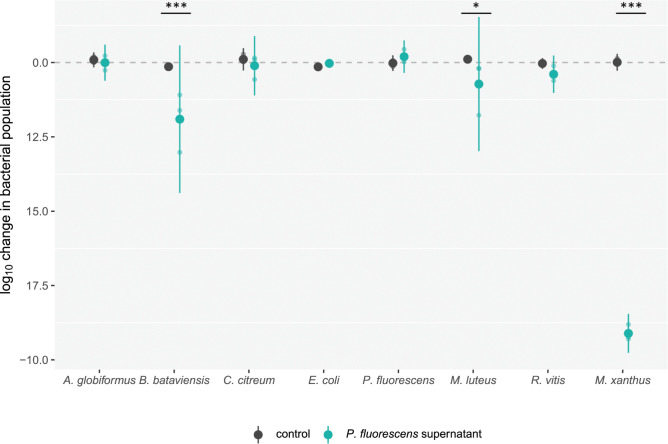
Ecological variation influences the character of many biotic interactions, but examples of predator-prey reversal mediated by abiotic context are few. We show that the temperature at which prey grow before interacting with a bacterial predator can determine the very direction of predation, reversing predator and prey identities. While Pseudomonas fluorescens reared at 32°C was extensively killed by the generalist predator Myxococcus xanthus, P. fluorescens reared at 22°C became the predator, slaughtering M. xanthus to extinction and growing on its remains. Beyond M. xanthus, diffusible molecules in P. fluorescens supernatant also killed 2 other phylogenetically distant species among several examined. Our results suggest that the sign of lethal microbial antagonisms may often change across abiotic gradients in natural microbial communities, with important ecological and evolutionary implications. They also suggest that a larger proportion of microbial warfare results in predation-the killing and consumption of organisms-than is generally recognized.
Copyright: © 2024 Vasse et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Dynamics of Solitary Predation by Myxococcus xanthus on Escherichia coli Observed at the Single-Cell Level.Appl Environ Microbiol. 2020 Jan 21;86(3):e02286-19. doi: 10.1128/AEM.02286-19. Print 2020 Jan 21. Appl Environ Microbiol. 2020. PMID: 31704687 Free PMC article.
-
Decomposing predation: testing for parameters that correlate with predatory performance by a social bacterium.Microb Ecol. 2013 Feb;65(2):415-23. doi: 10.1007/s00248-012-0135-6. Epub 2012 Nov 27. Microb Ecol. 2013. PMID: 23184156 Free PMC article.
-
The Genetics of Prey Susceptibility to Myxobacterial Predation: A Review, Including an Investigation into Pseudomonas aeruginosa Mutations Affecting Predation by Myxococcus xanthus.Microb Physiol. 2021;31(2):57-66. doi: 10.1159/000515546. Epub 2021 Apr 1. Microb Physiol. 2021. PMID: 33794538 Review.
-
Transcriptional changes when Myxococcus xanthus preys on Escherichia coli suggest myxobacterial predators are constitutively toxic but regulate their feeding.Microb Genom. 2018 Feb;4(2):e000152. doi: 10.1099/mgen.0.000152. Epub 2018 Jan 18. Microb Genom. 2018. PMID: 29345219 Free PMC article.
-
Myxococcus xanthus predation: an updated overview.Front Microbiol. 2024 Jan 24;15:1339696. doi: 10.3389/fmicb.2024.1339696. eCollection 2024. Front Microbiol. 2024. PMID: 38328431 Free PMC article. Review.
Cited by
-
Siderophores and competition for iron govern myxobacterial predation dynamics.ISME J. 2024 Jan 8;18(1):wrae077. doi: 10.1093/ismejo/wrae077. ISME J. 2024. PMID: 38696719 Free PMC article.
References
-
- Dunson WA, Travis J. The Role of Abiotic Factors in Community Organization. Am Nat. 1991;138(5):1067–1091.
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
