

The evolutionary history of human spindle genes includes back-and-forth gene flow with Neandertals
- PMID: 35816093
- PMCID: PMC9273211
- DOI: 10.7554/eLife.75464
The evolutionary history of human spindle genes includes back-and-forth gene flow with Neandertals
Abstract
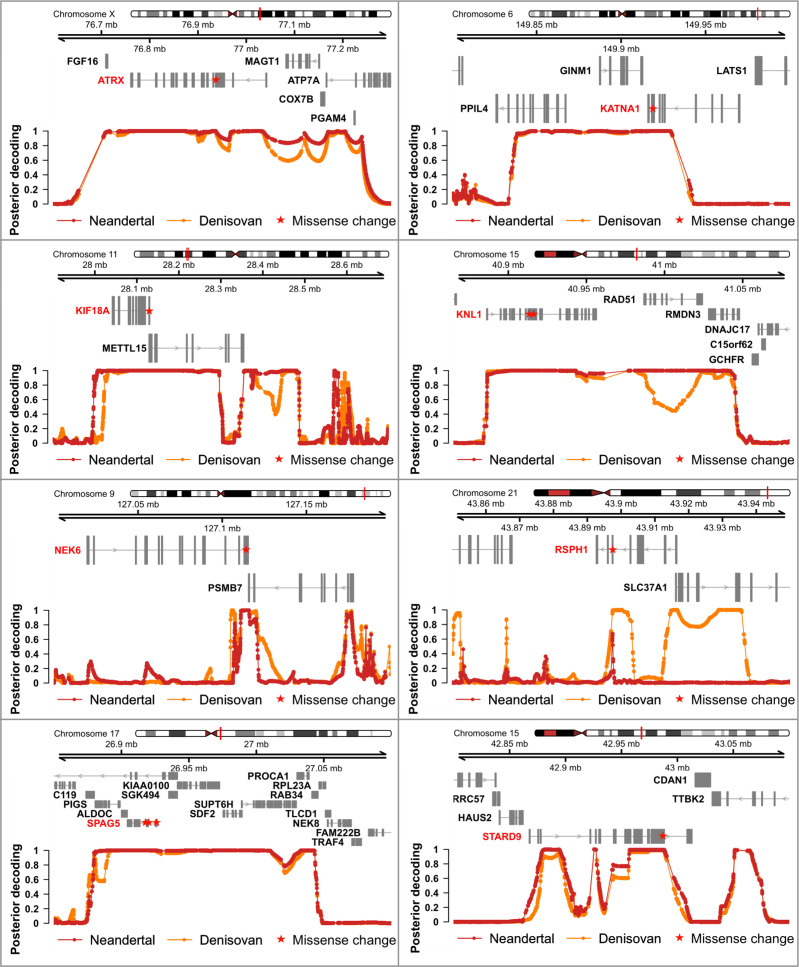
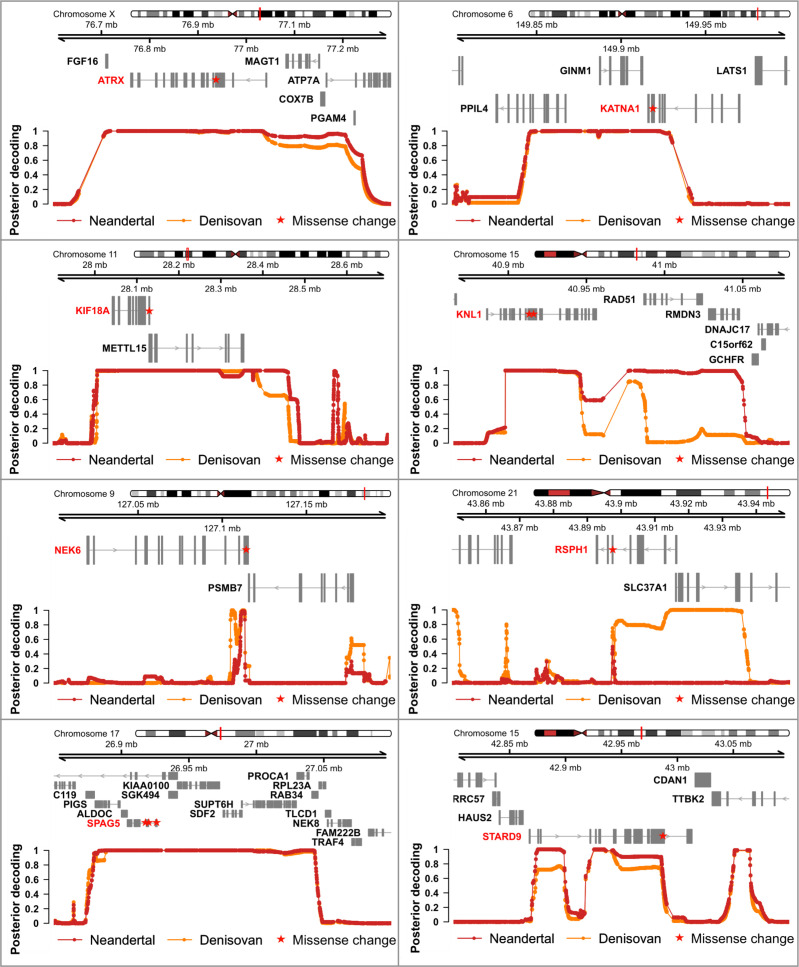
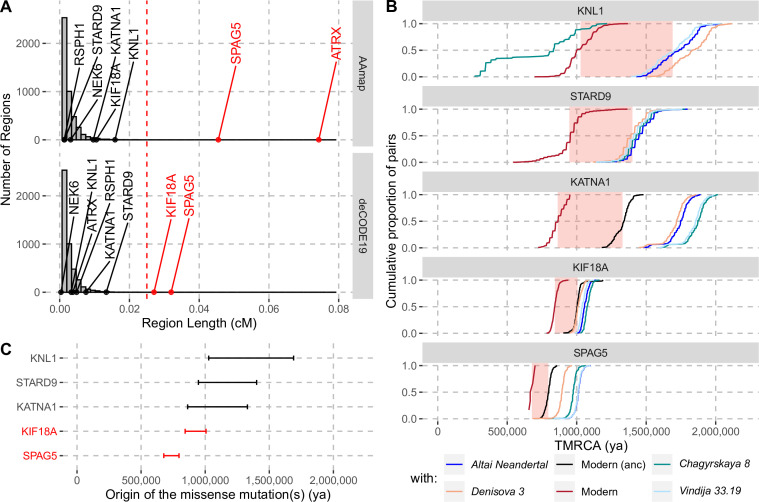
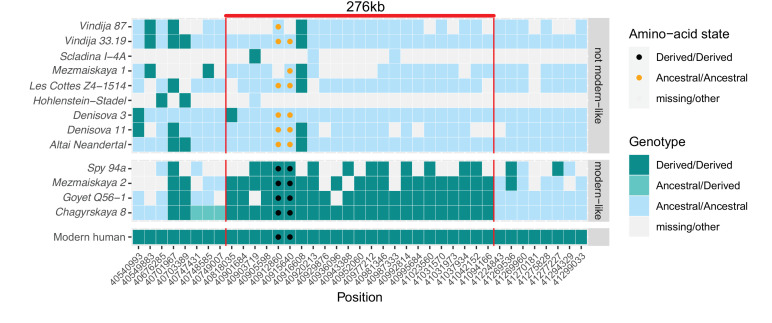
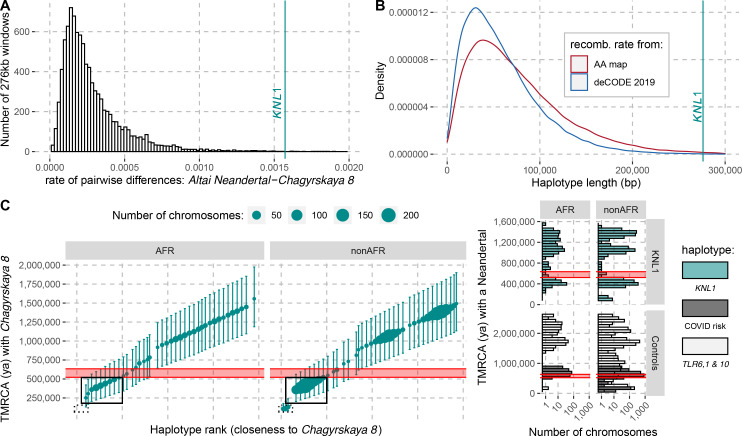
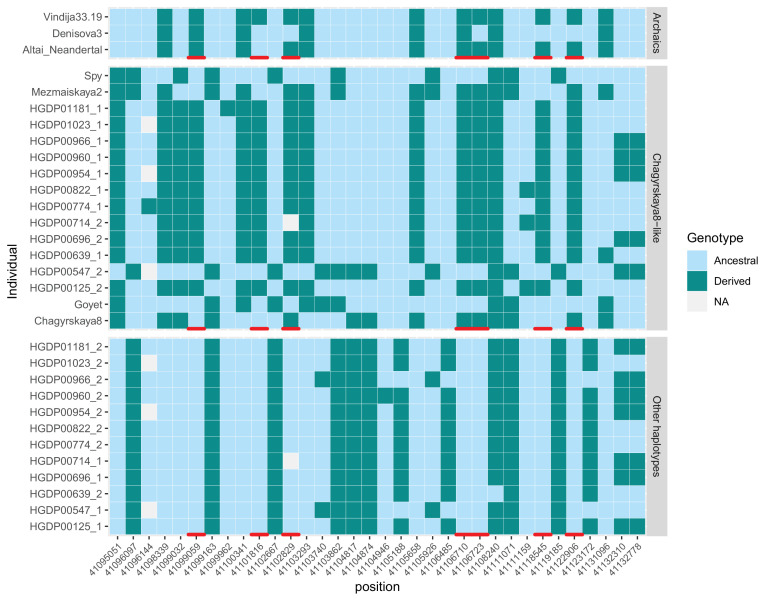
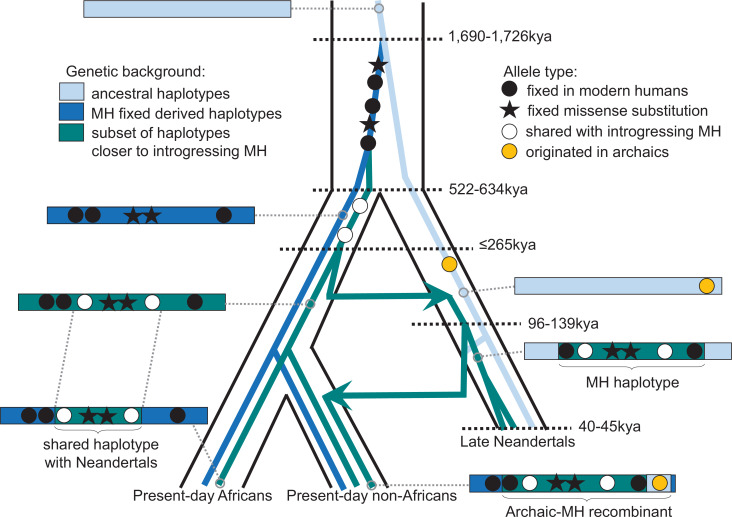
Proteins associated with the spindle apparatus, a cytoskeletal structure that ensures the proper segregation of chromosomes during cell division, experienced an unusual number of amino acid substitutions in modern humans after the split from the ancestors of Neandertals and Denisovans. Here, we analyze the history of these substitutions and show that some of the genes in which they occur may have been targets of positive selection. We also find that the two changes in the kinetochore scaffold 1 (KNL1) protein, previously believed to be specific to modern humans, were present in some Neandertals. We show that the KNL1 gene of these Neandertals shared a common ancestor with present-day Africans about 200,000 years ago due to gene flow from the ancestors (or relatives) of modern humans into Neandertals. Subsequently, some non-Africans inherited this modern human-like gene variant from Neandertals, but none inherited the ancestral gene variants. These results add to the growing evidence of early contacts between modern humans and archaic groups in Eurasia and illustrate the intricate relationships among these groups.
Keywords: evolutionary biology; gene flow; genetics; genomics; human; human evolution; neandertals; positive selection; spindle.
© 2022, Peyrégne et al.
Conflict of interest statement
SP, JK, BP, SP No competing interests declared
Figures
Similar articles
-
The date of interbreeding between Neandertals and modern humans.PLoS Genet. 2012;8(10):e1002947. doi: 10.1371/journal.pgen.1002947. Epub 2012 Oct 4. PLoS Genet. 2012. PMID: 23055938 Free PMC article.
-
IBD Sharing between Africans, Neandertals, and Denisovans.Genome Biol Evol. 2016 Dec 1;8(12):3406-3416. doi: 10.1093/gbe/evw234. Genome Biol Evol. 2016. PMID: 28158547 Free PMC article.
-
[Progresses on Neandertal genomics].Yi Chuan. 2012 Jun;34(6):659-65. doi: 10.3724/sp.j.1005.2012.00659. Yi Chuan. 2012. PMID: 22698735 Review. Chinese.
-
A draft sequence of the Neandertal genome.Science. 2010 May 7;328(5979):710-722. doi: 10.1126/science.1188021. Science. 2010. PMID: 20448178 Free PMC article.
-
The genetic changes that shaped Neandertals, Denisovans, and modern humans.Cell. 2024 Feb 29;187(5):1047-1058. doi: 10.1016/j.cell.2023.12.029. Epub 2024 Feb 16. Cell. 2024. PMID: 38367615 Review.
Cited by
-
More than a decade of genetic research on the Denisovans.Nat Rev Genet. 2024 Feb;25(2):83-103. doi: 10.1038/s41576-023-00643-4. Epub 2023 Sep 18. Nat Rev Genet. 2024. PMID: 37723347 Review.
-
Human genetic adaptation related to cellular zinc homeostasis.PLoS Genet. 2023 Sep 25;19(9):e1010950. doi: 10.1371/journal.pgen.1010950. eCollection 2023 Sep. PLoS Genet. 2023. PMID: 37747921 Free PMC article.
-
The Persian plateau served as hub for Homo sapiens after the main out of Africa dispersal.Nat Commun. 2024 Mar 25;15(1):1882. doi: 10.1038/s41467-024-46161-7. Nat Commun. 2024. PMID: 38528002 Free PMC article.
-
Evolution of Homo in the Middle and Late Pleistocene.J Hum Evol. 2022 Dec;173:103279. doi: 10.1016/j.jhevol.2022.103279. Epub 2022 Nov 11. J Hum Evol. 2022. PMID: 36375244 Free PMC article. Review.
-
The MUC19 gene in Denisovans, Neanderthals, and Modern Humans: An Evolutionary History of Recurrent Introgression and Natural Selection.bioRxiv [Preprint]. 2024 Dec 11:2023.09.25.559202. doi: 10.1101/2023.09.25.559202. bioRxiv. 2024. PMID: 37808839 Free PMC article. Preprint.
References
-
- Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, Harris MA, Hill DP, Issel-Tarver L, Kasarskis A, Lewis S, Matese JC, Richardson JE, Ringwald M, Rubin GM, Sherlock G. Gene Ontology: tool for the unification of biology. Nature Genetics. 2000;25:25–29. doi: 10.1038/75556. - DOI - PMC - PubMed
-
- Bergström A, McCarthy SA, Hui R, Almarri MA, Ayub Q, Danecek P, Chen Y, Felkel S, Hallast P, Kamm J, Blanché H, Deleuze J-F, Cann H, Mallick S, Reich D, Sandhu MS, Skoglund P, Scally A, Xue Y, Durbin R, Tyler-Smith C. Insights into human genetic variation and population history from 929 diverse genomes. Science (New York, N.Y.) 2020;367:eaay5012. doi: 10.1126/science.aay5012. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
