

Kin discrimination drives territorial exclusion during Bacillus subtilis swarming and restrains exploitation of surfactin
- PMID: 34650232
- PMCID: PMC8857193
- DOI: 10.1038/s41396-021-01124-4
Kin discrimination drives territorial exclusion during Bacillus subtilis swarming and restrains exploitation of surfactin
Abstract
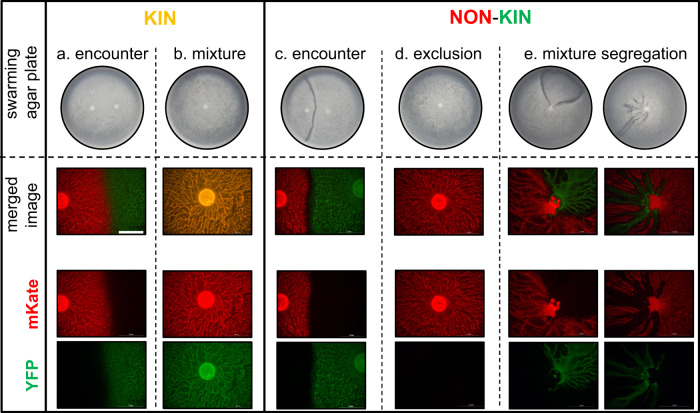
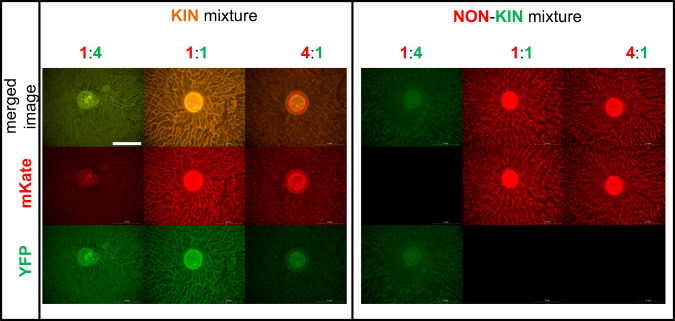
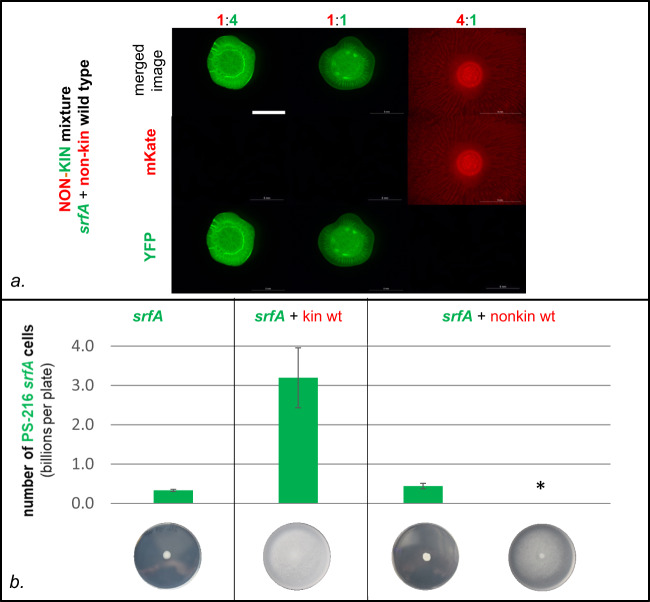
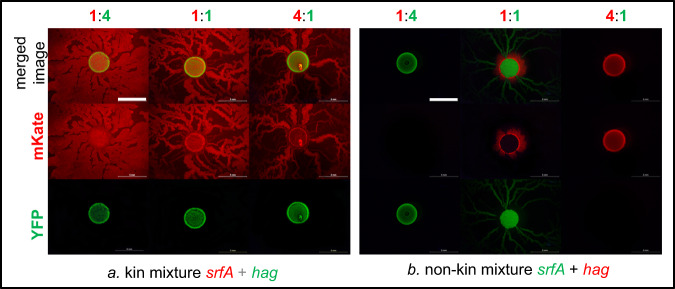
Swarming is the collective movement of bacteria across a surface. It requires the production of surfactants (public goods) to overcome surface tension and provides an excellent model to investigate bacterial cooperation. Previously, we correlated swarm interaction phenotypes with kin discrimination between B. subtilis soil isolates, by showing that less related strains form boundaries between swarms and highly related strains merge. However, how kin discrimination affects cooperation and territoriality in swarming bacteria remains little explored. Here we show that the pattern of surface colonization by swarming mixtures is influenced by kin types. Closely related strain mixtures colonize the surface in a mixed swarm, while mixtures of less related strains show competitive exclusion as only one strain colonizes the surface. The outcome of nonkin swarm expansion depends on the initial ratio of the competing strains, indicating positive frequency-dependent competition. We find that addition of surfactin (a public good excreted from cells) can complement the swarming defect of nonkin mutants, whereas close encounters in nonkin mixtures lead to territorial exclusion, which limits the exploitation of surfactin by nonkin nonproducers. The work suggests that kin discrimination driven competitive territorial exclusion may be an important determinant for the success of cooperative surface colonization.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Kin Discrimination Modifies Strain Distribution, Spatial Segregation, and Incorporation of Extracellular Matrix Polysaccharide Mutants of Bacillus subtilis Strains into Mixed Floating Biofilms.Appl Environ Microbiol. 2022 Sep 22;88(18):e0087122. doi: 10.1128/aem.00871-22. Epub 2022 Sep 12. Appl Environ Microbiol. 2022. PMID: 36094206 Free PMC article.
-
Nonkin interactions between Bacillus subtilis soil isolates limit the spread of swarming deficient cheats.ISME J. 2024 Jan 8;18(1):wrae199. doi: 10.1093/ismejo/wrae199. ISME J. 2024. PMID: 39375016 Free PMC article.
-
Bacillus subtilis Protects Public Goods by Extending Kin Discrimination to Closely Related Species.mBio. 2017 Jul 5;8(4):e00723-17. doi: 10.1128/mBio.00723-17. mBio. 2017. PMID: 28679746 Free PMC article.
-
Social behaviours by Bacillus subtilis: quorum sensing, kin discrimination and beyond.Mol Microbiol. 2018 Dec;110(6):863-878. doi: 10.1111/mmi.14127. Epub 2018 Nov 1. Mol Microbiol. 2018. PMID: 30218468 Free PMC article. Review.
-
Kin Recognition in Bacteria.Annu Rev Microbiol. 2016 Sep 8;70:143-60. doi: 10.1146/annurev-micro-102215-095325. Epub 2016 Jun 17. Annu Rev Microbiol. 2016. PMID: 27359217 Free PMC article. Review.
Cited by
-
Bacillus subtilis extracellular protease production incurs a context-dependent cost.Mol Microbiol. 2023 Aug;120(2):105-121. doi: 10.1111/mmi.15110. Epub 2023 Jun 28. Mol Microbiol. 2023. PMID: 37380434 Free PMC article.
-
Involvement of Flagellin in Kin Recognition between Bacillus velezensis Strains.mSystems. 2022 Dec 20;7(6):e0077822. doi: 10.1128/msystems.00778-22. Epub 2022 Oct 11. mSystems. 2022. PMID: 36218362 Free PMC article.
-
EAM highlights in FEMS 2023: from the Petri dish to planet Earth.Microlife. 2023 Nov 3;4:uqad045. doi: 10.1093/femsml/uqad045. eCollection 2023. Microlife. 2023. PMID: 38107236 Free PMC article.
-
Kin Discrimination Modifies Strain Distribution, Spatial Segregation, and Incorporation of Extracellular Matrix Polysaccharide Mutants of Bacillus subtilis Strains into Mixed Floating Biofilms.Appl Environ Microbiol. 2022 Sep 22;88(18):e0087122. doi: 10.1128/aem.00871-22. Epub 2022 Sep 12. Appl Environ Microbiol. 2022. PMID: 36094206 Free PMC article.
-
Annulment of Bacterial Antagonism Improves Plant Beneficial Activity of a Bacillus velezensis Consortium.Appl Environ Microbiol. 2022 Apr 26;88(8):e0024022. doi: 10.1128/aem.00240-22. Epub 2022 Apr 5. Appl Environ Microbiol. 2022. PMID: 35380452 Free PMC article.
References
-
- Claessen D, Rozen DE, Kuipers OP, Søgaard-Andersen L, van Wezel GP. Bacterial solutions to multicellularity: a tale of biofilms, filaments and fruiting bodies. Nat Rev Microbiol. 2014;12:115. - PubMed
-
- Dunny GM, Brickman TJ, Dworkin M. Multicellular behavior in bacteria: communication, cooperation, competition and cheating. BioEssays. 2008;30:296–8. - PubMed
-
- Fisher Roberta M, Cornwallis Charlie K, West SA. Group formation, relatedness, and the evolution of multicellularity. Curr Biol. 2013;23:1120–5. - PubMed
-
- Shapiro J. Bacteria as multicellular organisms. Sci Am. 1988;258:82–9. - PubMed
