

Understanding the Role of Anti-PEG Antibodies in the Complement Activation by Doxil in Vitro
- PMID: 30002298
- PMCID: PMC6100003
- DOI: 10.3390/molecules23071700
Understanding the Role of Anti-PEG Antibodies in the Complement Activation by Doxil in Vitro
Abstract
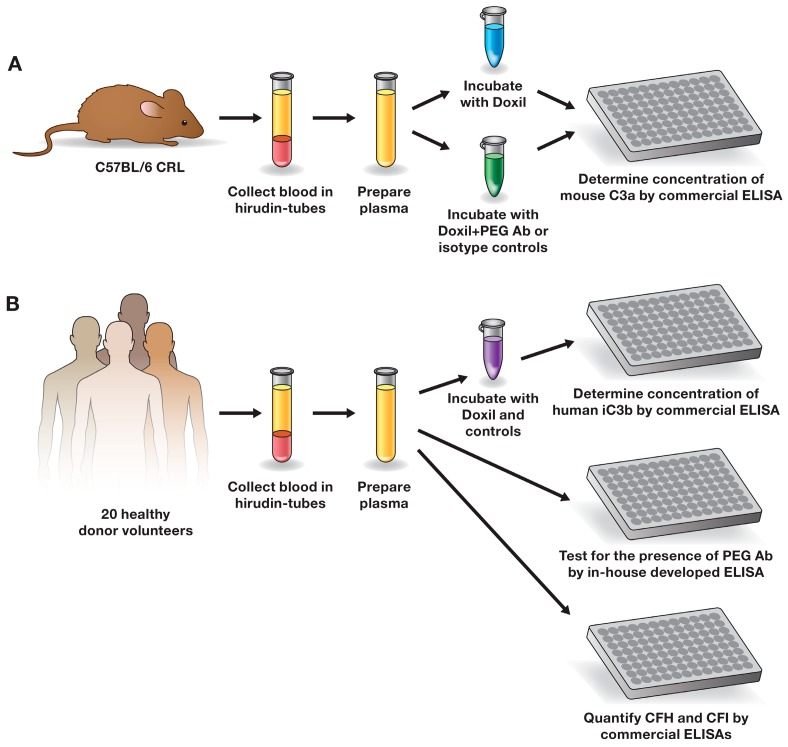
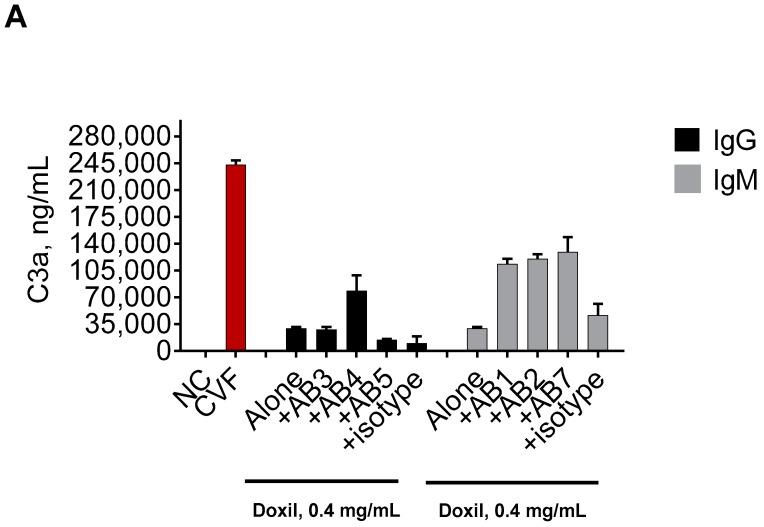
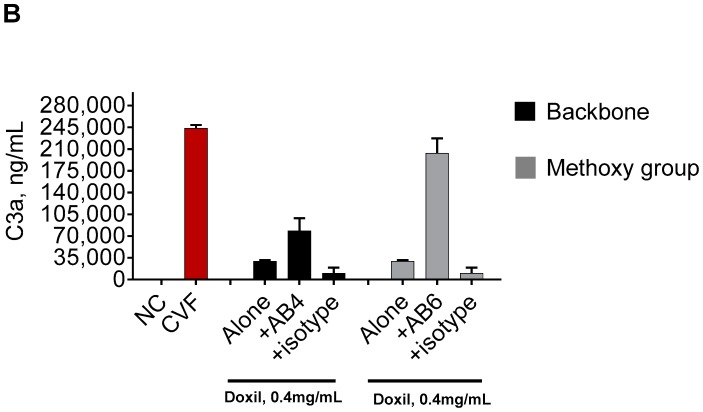
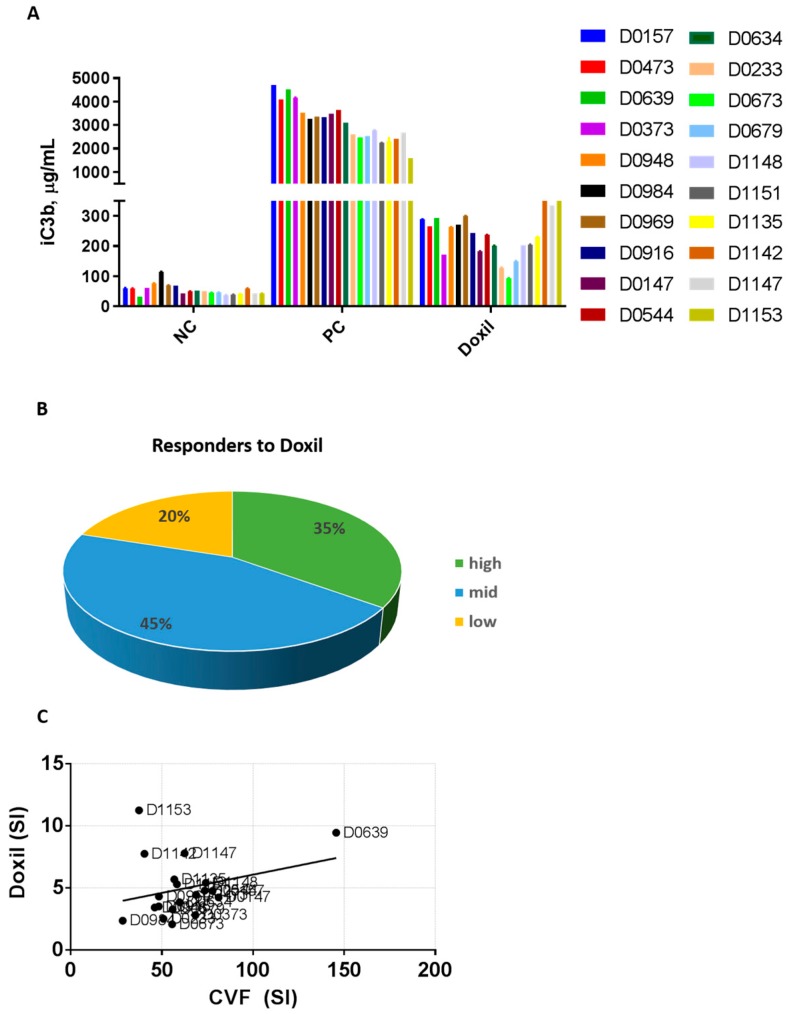
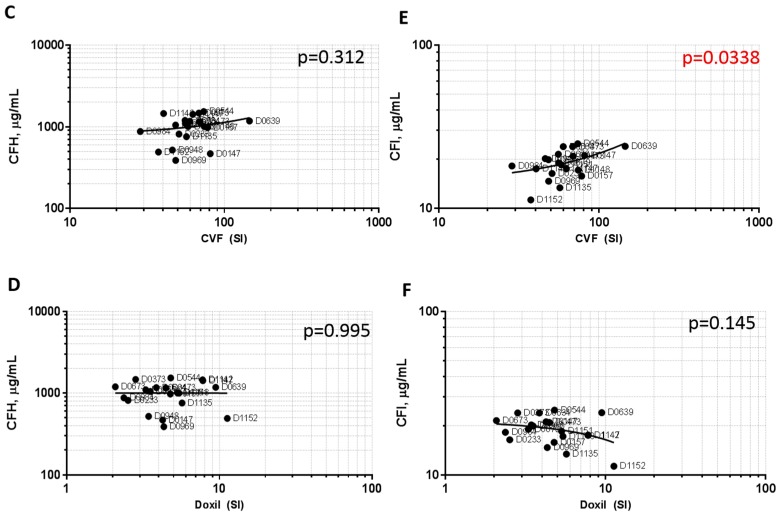
Infusion reactions (IRs) are common immune-mediated side effects in patients treated with a variety of drug products, including, but not limited to, nanotechnology formulations. The mechanism of IRs is not fully understood. One of the best studied mechanisms of IRs to nanomedicines is the complement activation. However, it is largely unknown why some patients develop reactions to nanomedicines while others do not, and why some nanoparticles are more reactogenic than others. One of the theories is that the pre-existing anti-polyethylene glycol (PEG) antibodies initiate the complement activation and IRs in patients. In this study, we investigated this hypothesis in the case of PEGylated liposomal doxorubicin (Doxil), which, when used in a clinical setting, is known to induce IRs; referred to as complement activation-related pseudoallergy (CARPA) in sensitive individuals. We conducted the study in vitro using plasma derived from C57BL/6 mice and twenty human donor volunteers. We used mouse plasma to test a library of well-characterized mouse monoclonal antibodies with different specificity and affinity to PEG as it relates to the complement activation by Doxil. We determined the levels of pre-existing polyclonal antibodies that bind to PEG, methoxy-PEG, and PEGylated liposomes in human plasma, and we also assessed complement activation by Doxil and concentrations of complement inhibitory factors H and I in these human plasma specimens. The affinity, specificity, and other characteristics of the human polyclonal antibodies are not known at this time. Our data demonstrate that under in vitro conditions, some anti-PEG antibodies contribute to the complement activation by Doxil. Such contribution, however, needs to be considered in the context of other factors, including, but not limited to, antibody class, type, clonality, epitope specificity, affinity, and titer. In addition, our data contribute to the knowledge base used to understand and improve nanomedicine safety.
Keywords: CARPA; anti-PEG antibody; complement activation; hypersensitivity; immunotoxicity; infusion reaction; liposomes; nanoparticles.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Features of complement activation-related pseudoallergy to liposomes with different surface charge and PEGylation: comparison of the porcine and rat responses.J Control Release. 2014 Dec 10;195:2-10. doi: 10.1016/j.jconrel.2014.08.009. Epub 2014 Aug 19. J Control Release. 2014. PMID: 25148822
-
Doxebo (doxorubicin-free Doxil-like liposomes) is safe to use as a pre-treatment to prevent infusion reactions to PEGylated nanodrugs.J Control Release. 2019 Jul 28;306:138-148. doi: 10.1016/j.jconrel.2019.06.007. Epub 2019 Jun 6. J Control Release. 2019. PMID: 31176656
-
Activation of complement by therapeutic liposomes and other lipid excipient-based therapeutic products: prediction and prevention.Adv Drug Deliv Rev. 2011 Sep 16;63(12):1020-30. doi: 10.1016/j.addr.2011.06.017. Epub 2011 Jul 14. Adv Drug Deliv Rev. 2011. PMID: 21787819 Review.
-
Prevention of infusion reactions to PEGylated liposomal doxorubicin via tachyphylaxis induction by placebo vesicles: a porcine model.J Control Release. 2012 Jun 10;160(2):382-7. doi: 10.1016/j.jconrel.2012.02.029. Epub 2012 Mar 6. J Control Release. 2012. PMID: 22421426
-
A porcine model of complement-mediated infusion reactions to drug carrier nanosystems and other medicines.Adv Drug Deliv Rev. 2012 Dec;64(15):1706-16. doi: 10.1016/j.addr.2012.07.005. Epub 2012 Jul 20. Adv Drug Deliv Rev. 2012. PMID: 22820530 Review.
Cited by
-
Clinical Relevance of Pre-Existing and Treatment-Induced Anti-Poly(Ethylene Glycol) Antibodies.Regen Eng Transl Med. 2022;8(1):32-42. doi: 10.1007/s40883-021-00198-y. Epub 2021 Mar 25. Regen Eng Transl Med. 2022. PMID: 33786367 Free PMC article. Review.
-
The conundrum of COVID-19 mRNA vaccine-induced anaphylaxis.J Allergy Clin Immunol Glob. 2023 Feb;2(1):1-13. doi: 10.1016/j.jacig.2022.10.003. Epub 2022 Dec 13. J Allergy Clin Immunol Glob. 2023. PMID: 36532656 Free PMC article. Review.
-
Impact of PEG sensitization on the efficacy of PEG hydrogel-mediated tissue engineering.Nat Commun. 2024 Apr 18;15(1):3283. doi: 10.1038/s41467-024-46327-3. Nat Commun. 2024. PMID: 38637507 Free PMC article.
-
PEG Conjugated Zein Nanoparticles for In Vivo Use.Pharmaceutics. 2022 Aug 31;14(9):1831. doi: 10.3390/pharmaceutics14091831. Pharmaceutics. 2022. PMID: 36145579 Free PMC article.
-
Revisiting the outstanding questions in cancer nanomedicine with a future outlook.Nanoscale Adv. 2021 Dec 22;4(3):634-653. doi: 10.1039/d1na00810b. eCollection 2022 Feb 1. Nanoscale Adv. 2021. PMID: 36131837 Free PMC article. Review.
References
-
- Corominas M., Gastaminza G., Lobera T. Hypersensitivity reactions to biological drugs. J. Investig. Allergol. Clin. Immunol. 2014;24:212–225. - PubMed
-
- Johansson S.G.O., Bieber T., Dahl R., Friedmann P.S., Lanier B.Q., Lockey R.F., Motala C., Martell J.A.O., Platts-Mills T.A., Ring J., et al. Revised nomenclature for allergy for global use: Report of the Nomenclature Review Committee of the World Allergy Organization, October 2003. J. Allergy Clin. Immunol. 2004;113:832–836. doi: 10.1016/j.jaci.2003.12.591. - DOI - PubMed
-
- Johansson S.G.O., Hourihane J.B., Bousquet J., Bruijnzeel-Koomen C., Dreborg S., Haahtela T., Kowalski M.L., Mygind N., Ring J., van Cauwenberge P., et al. A revised nomenclature for allergy: An EAACI position statement from the EAACI nomenclature task force. Allergy. 2001;56:813–824. doi: 10.1034/j.1398-9995.2001.t01-1-00001.x. - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
