

Effects of prey, pitcher age, and microbes on acid phosphatase activity in fluid from pitchers of Sarracenia purpurea (Sarraceniaceae)
- PMID: 28719666
- PMCID: PMC5515422
- DOI: 10.1371/journal.pone.0181252
Effects of prey, pitcher age, and microbes on acid phosphatase activity in fluid from pitchers of Sarracenia purpurea (Sarraceniaceae)
Abstract
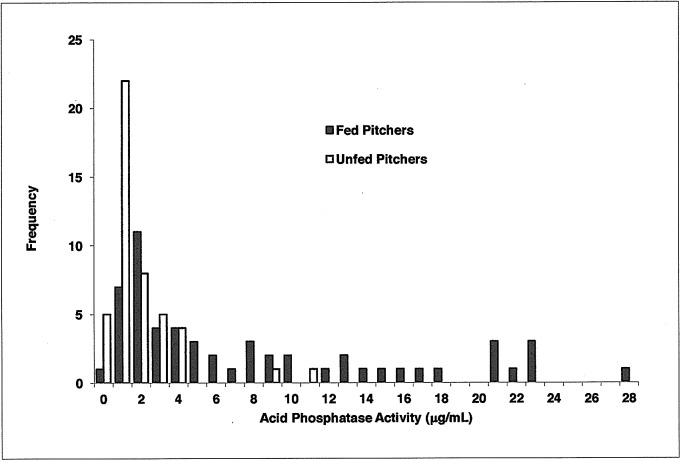
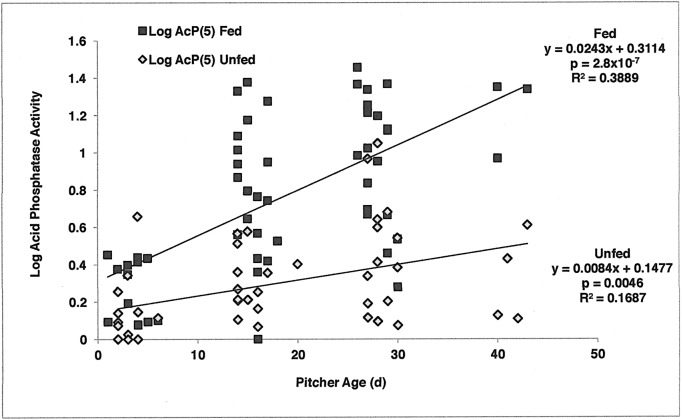
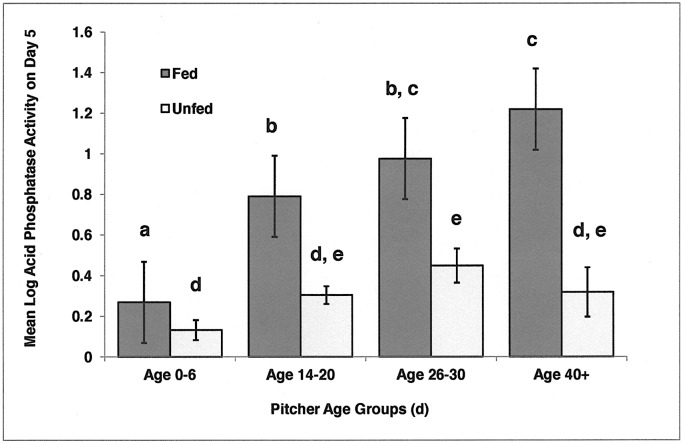
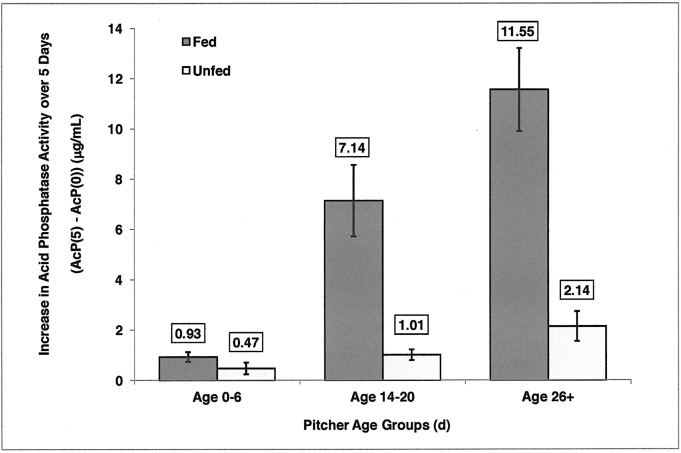
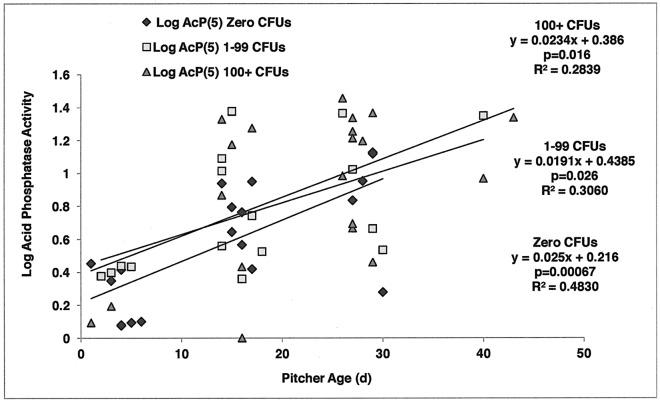
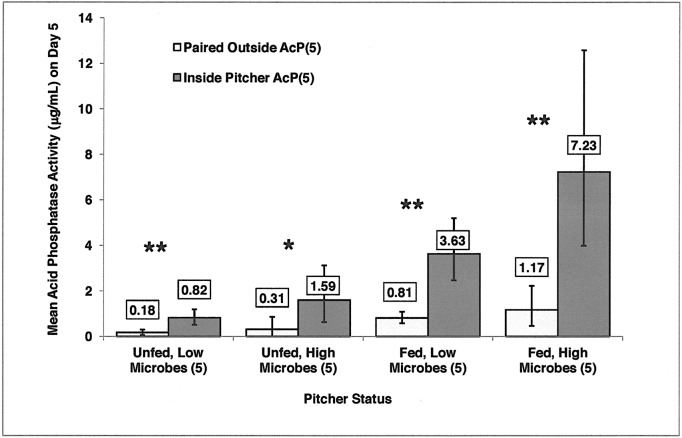
Carnivory in pitcher plants generally involves digestion of prey, by the plant itself, by symbionts, or both. While symbionts appear to be important in the digestion of prey in Sarracenia purpurea, the importance of pitcher-derived enzymes is less well documented. Our goal was to reduce microbial numbers in pitcher fluid in order to measure the acid phosphatase activity attributable to the pitchers themselves. Preliminary experiments indicated that various antibiotics were minimally effective at reducing microbial populations and that antibiotic-resistant microbes were easily cultured from pitcher fluid. Consequently, we measured the abundance of culturable microbes in every sample taken for the measurement of acid phosphatase activity. Pitchers fed with one sterilized ant had higher levels of acid phosphatase activity than unfed pitchers. Older pitchers were more responsive to feeding than young pitchers. Pitchers with high levels of microbes (on Day 5) had higher acid phosphatase activity than pitchers with low levels of microbes. However, fed pitchers were not more likely to have higher microbe levels and microbe levels were not related to pitcher age. When fluid samples from inside the pitcher were compared to appropriate controls incubated outside the pitcher, acid phosphatase activity was higher inside the pitcher. Results from the feeding experiments are consistent with a primary role of microbes in the digestion of prey in pitchers of S. purpurea. However, the relationship between pitcher age and enzyme activity is not a function of microbes in the pitcher fluid and may depend on enzymes produced by the plant. Our methods would not detect microbes embedded on the inner surface of the pitcher; and if they survived the alcohol rinse and antibiotics, we cannot rule out microbes as the source of the relationship between pitcher age and acid phosphatase activity.
Conflict of interest statement
Figures
Similar articles
-
Selective Bacterial Community Enrichment between the Pitcher Plants Sarracenia minor and Sarracenia flava.Microbiol Spectr. 2021 Dec 22;9(3):e0069621. doi: 10.1128/Spectrum.00696-21. Epub 2021 Nov 24. Microbiol Spectr. 2021. PMID: 34817222 Free PMC article.
-
Detection of methanogenic archaea in the pitchers of the Northern pitcher plant (Sarracenia purpurea ).Can J Microbiol. 2012 Feb;58(2):189-94. doi: 10.1139/w11-117. Epub 2012 Jan 20. Can J Microbiol. 2012. PMID: 22263906
-
Efficiency of insect capture by Sarracenia purpurea (Sarraceniaceae), the northern pitcher plant.Am J Bot. 1998 Jan;85(1):88. Am J Bot. 1998. PMID: 21684883
-
Traps of carnivorous pitcher plants as a habitat: composition of the fluid, biodiversity and mutualistic activities.Ann Bot. 2011 Feb;107(2):181-94. doi: 10.1093/aob/mcq238. Epub 2010 Dec 15. Ann Bot. 2011. PMID: 21159782 Free PMC article. Review.
-
Carnivorous pitcher plants: insights in an old topic.Phytochemistry. 2011 Sep;72(13):1678-82. doi: 10.1016/j.phytochem.2010.11.024. Epub 2010 Dec 22. Phytochemistry. 2011. PMID: 21185041 Review.
Cited by
-
Bacterial Recruitment to Carnivorous Pitcher Plant Communities: Identifying Sources Influencing Plant Microbiome Composition and Function.Front Microbiol. 2022 Mar 14;13:791079. doi: 10.3389/fmicb.2022.791079. eCollection 2022. Front Microbiol. 2022. PMID: 35359741 Free PMC article.
-
Selective Bacterial Community Enrichment between the Pitcher Plants Sarracenia minor and Sarracenia flava.Microbiol Spectr. 2021 Dec 22;9(3):e0069621. doi: 10.1128/Spectrum.00696-21. Epub 2021 Nov 24. Microbiol Spectr. 2021. PMID: 34817222 Free PMC article.
-
Characterization and Comparison of Convergence Among Cephalotus follicularis Pitcher Plant-Associated Communities With Those of Nepenthes and Sarracenia Found Worldwide.Front Plant Sci. 2022 Jun 6;13:887635. doi: 10.3389/fpls.2022.887635. eCollection 2022. Front Plant Sci. 2022. PMID: 35734258 Free PMC article.
-
An inquiline mosquito modulates microbial diversity and function in an aquatic microecosystem.Mol Ecol. 2024 Apr;33(7):e17314. doi: 10.1111/mec.17314. Epub 2024 Mar 5. Mol Ecol. 2024. PMID: 38441172
-
Discovery of digestive enzymes in carnivorous plants with focus on proteases.PeerJ. 2018 Jun 5;6:e4914. doi: 10.7717/peerj.4914. eCollection 2018. PeerJ. 2018. PMID: 29888132 Free PMC article.
References
-
- Chase MW, Christenhusz MJM, Sanders D, Fay MF. Murderous plants: Victorian Gothic, Darwin and modern insights into vegetable carnivory. Bot J Linn Soc. 2009; 161: 329–356.
-
- Ellison AM, Gotelli NJ. Energetics and the evolution of carnivorous plants–Darwin’s ‘most wonderful plants in the world’. J Exp Bot. 2009; 60: 19–42. doi: 10.1093/jxb/ern179 - DOI - PubMed
-
- Mithöfer A. Carnivorous pitcher plants: Insights into an old topic. Phytochemistry. 2011; 13: 1678–1682. - PubMed
-
- Anderson B, Midgley JJ. Digestive mutualism, an alternate pathway in plant carnivory. Oikos. 2003; 102: 221–224.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
