

ARL3 regulates trafficking of prenylated phototransduction proteins to the rod outer segment
- PMID: 26936825
- PMCID: PMC5062590
- DOI: 10.1093/hmg/ddw077
ARL3 regulates trafficking of prenylated phototransduction proteins to the rod outer segment
Abstract
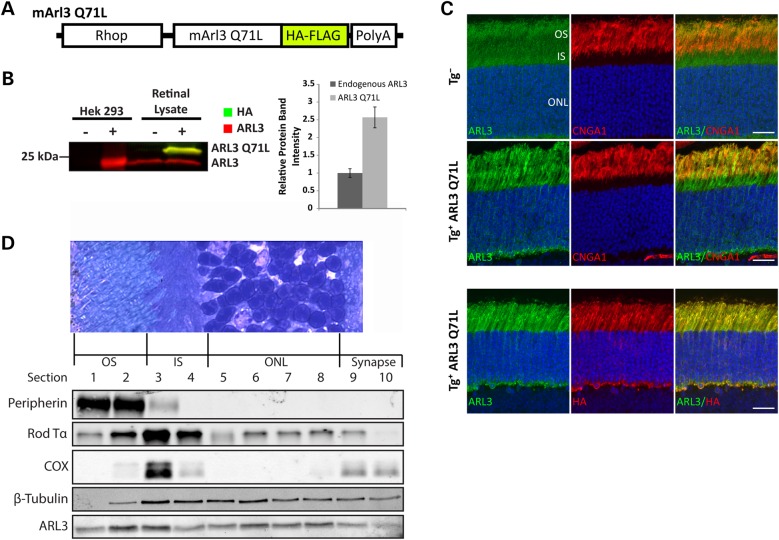
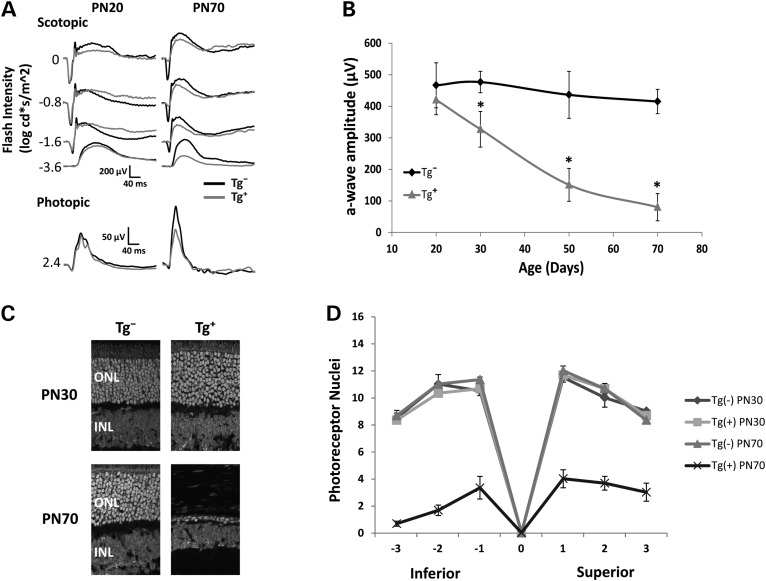
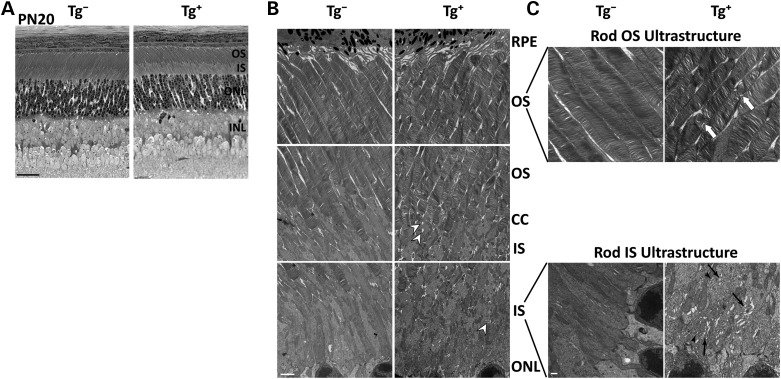
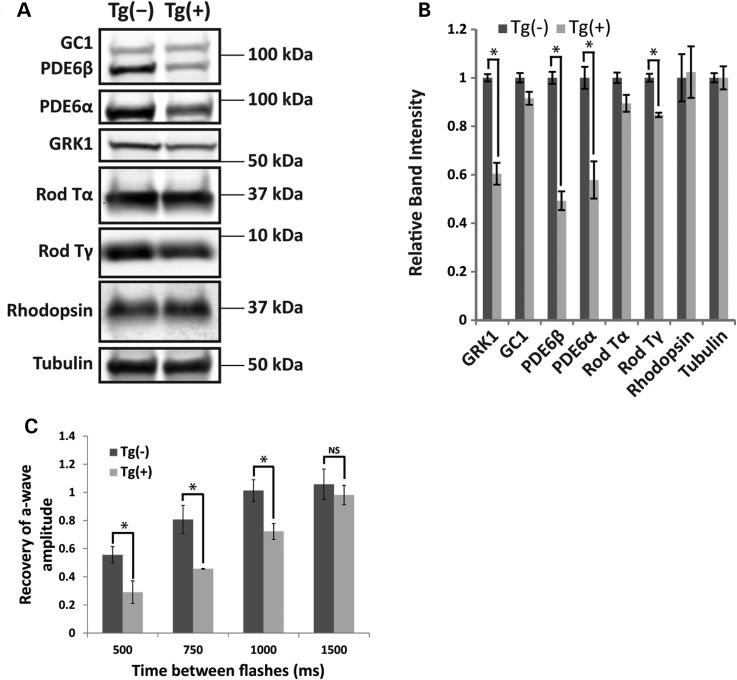
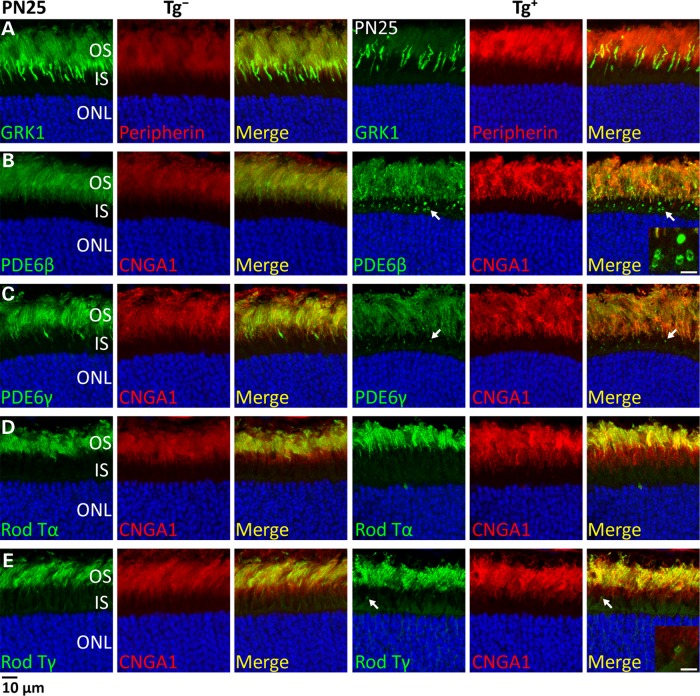
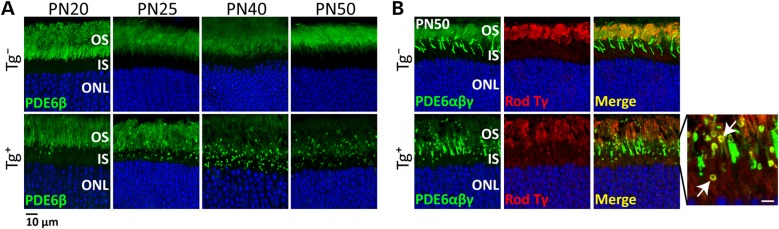
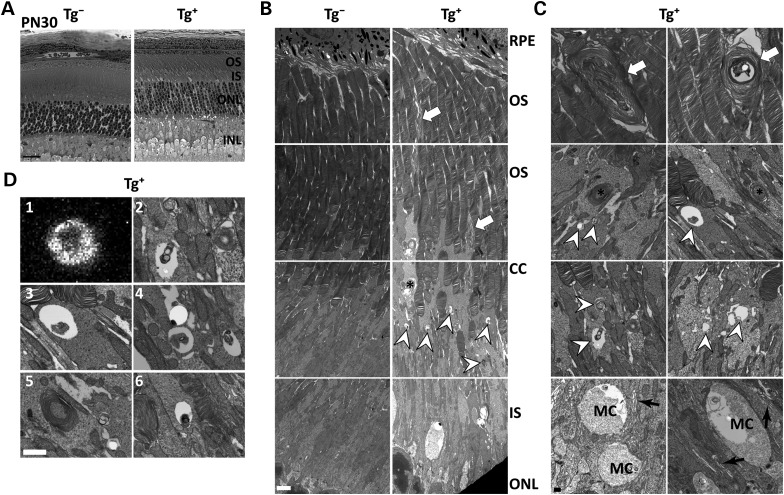
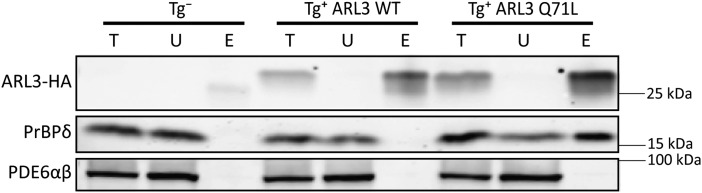
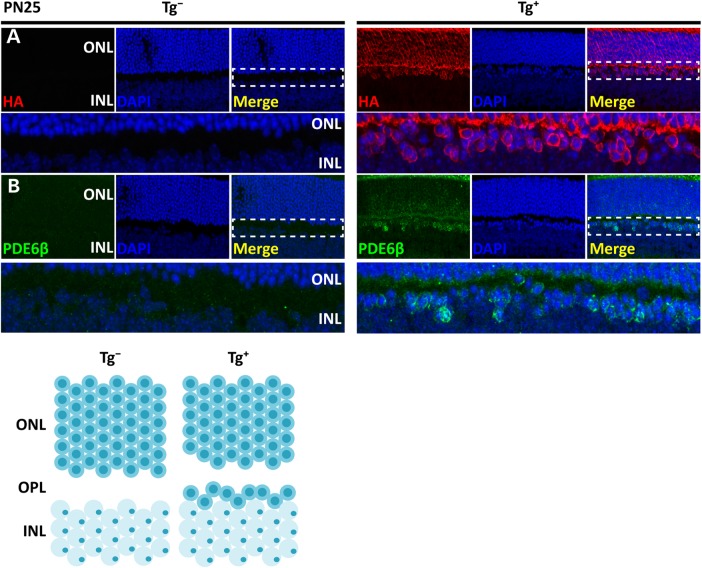
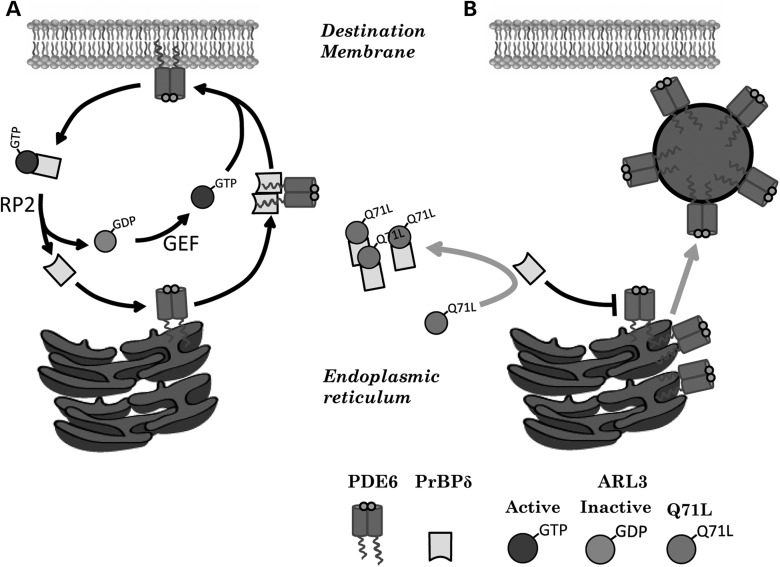
The small GTPase, ADP-ribosylation factor-like 3 (ARL3), has been proposed to participate in the transport of proteins in photoreceptor cells. Moreover, it has been implicated in the pathogenesis associated with X-linked retinitis pigmentosa (XLRP) resulting from mutations in the ARL3 GTPase activating protein, retinitis pigmentosa 2 (RP2). To determine the importance of ARL3 in rod photoreceptor cells, we generated transgenic mice expressing a dominant active form of ARL3 (ARL3-Q71L) under a rod-specific promoter. ARL3-Q71L animals exhibited extensive rod cell death after post-natal day 30 (PN30) and degeneration was complete by PN70. Prior to the onset of cell death, rod photoresponse was significantly reduced along with a robust decrease in rod phosphodiesterase 6 (PDE6) and G-protein receptor kinase-1 (GRK1) levels. Furthermore, assembled phosphodiesterase-6 (PDE6) subunits, rod transducin and G-protein receptor kinase-1 (GRK1) accumulated on large punctate structures within the inner segment in ARL3-Q71L retina. Defective trafficking of prenylated proteins is likely due to sequestration of prenyl binding protein δ (PrBPδ) by ARL3-Q71L as we demonstrate a specific interaction between these proteins in the retina. Unexpectedly, our studies also revealed a novel role for ARL3 in the migration of photoreceptor nuclei. In conclusion, this study identifies ARL3 as a key player in prenylated protein trafficking in rod photoreceptor cells and establishes the potential role for ARL3 dysregulation in the pathogenesis of RP2-related forms of XLRP.
© The Author 2016. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
Similar articles
-
Pathogenic mutations in retinitis pigmentosa 2 predominantly result in loss of RP2 protein stability in humans and zebrafish.J Biol Chem. 2017 Apr 14;292(15):6225-6239. doi: 10.1074/jbc.M116.760314. Epub 2017 Feb 16. J Biol Chem. 2017. PMID: 28209709 Free PMC article.
-
Mistrafficking of prenylated proteins causes retinitis pigmentosa 2.FASEB J. 2015 Mar;29(3):932-42. doi: 10.1096/fj.14-257915. Epub 2014 Nov 24. FASEB J. 2015. PMID: 25422369 Free PMC article.
-
Depressing time: Waiting, melancholia, and the psychoanalytic practice of care.In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. PMID: 36137063 Free Books & Documents. Review.
-
In vivo photoreceptor base editing ameliorates rhodopsin-E150K autosomal-recessive retinitis pigmentosa in mice.Proc Natl Acad Sci U S A. 2024 Nov 26;121(48):e2416827121. doi: 10.1073/pnas.2416827121. Epub 2024 Nov 18. Proc Natl Acad Sci U S A. 2024. PMID: 39556729 Free PMC article.
-
Impact of residual disease as a prognostic factor for survival in women with advanced epithelial ovarian cancer after primary surgery.Cochrane Database Syst Rev. 2022 Sep 26;9(9):CD015048. doi: 10.1002/14651858.CD015048.pub2. Cochrane Database Syst Rev. 2022. PMID: 36161421 Free PMC article. Review.
Cited by
-
An intrinsic compartmentalization code for peripheral membrane proteins in photoreceptor neurons.J Cell Biol. 2019 Nov 4;218(11):3753-3772. doi: 10.1083/jcb.201906024. Epub 2019 Oct 8. J Cell Biol. 2019. PMID: 31594805 Free PMC article.
-
A Novel ARL3 Gene Mutation Associated With Autosomal Dominant Retinal Degeneration.Front Cell Dev Biol. 2021 Aug 17;9:720782. doi: 10.3389/fcell.2021.720782. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34485303 Free PMC article.
-
Disrupting the ciliary gradient of active Arl3 affects rod photoreceptor nuclear migration.Elife. 2023 Jan 4;12:e80533. doi: 10.7554/eLife.80533. Elife. 2023. PMID: 36598133 Free PMC article.
-
Functional compartmentalization of photoreceptor neurons.Pflugers Arch. 2021 Sep;473(9):1493-1516. doi: 10.1007/s00424-021-02558-7. Epub 2021 Apr 20. Pflugers Arch. 2021. PMID: 33880652 Free PMC article. Review.
-
Roles for ELMOD2 and Rootletin in ciliogenesis.Mol Biol Cell. 2021 Apr 15;32(8):800-822. doi: 10.1091/mbc.E20-10-0635. Epub 2021 Feb 17. Mol Biol Cell. 2021. PMID: 33596093 Free PMC article.
References
-
- Gillingham A.K., Munro S. (2007) The small G proteins of the Arf family and their regulators. Annu. Rev. Cell Dev. Biol., 23, 579–611. - PubMed
-
- Veltel S., Gasper R., Eisenacher E., Wittinghofer A. (2008) The retinitis pigmentosa 2 gene product is a GTPase-activating protein for Arf-like 3. Nat. Struct. Mol. Biol., 15, 373–380. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
