

An Essential Poison: Synthesis and Degradation of Cyclic Di-AMP in Bacillus subtilis
- PMID: 26240071
- PMCID: PMC4573722
- DOI: 10.1128/JB.00564-15
An Essential Poison: Synthesis and Degradation of Cyclic Di-AMP in Bacillus subtilis
Abstract
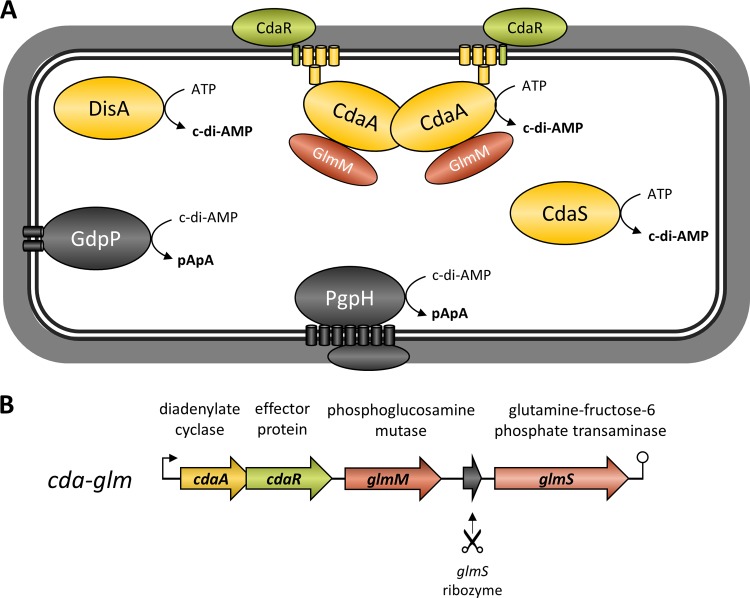
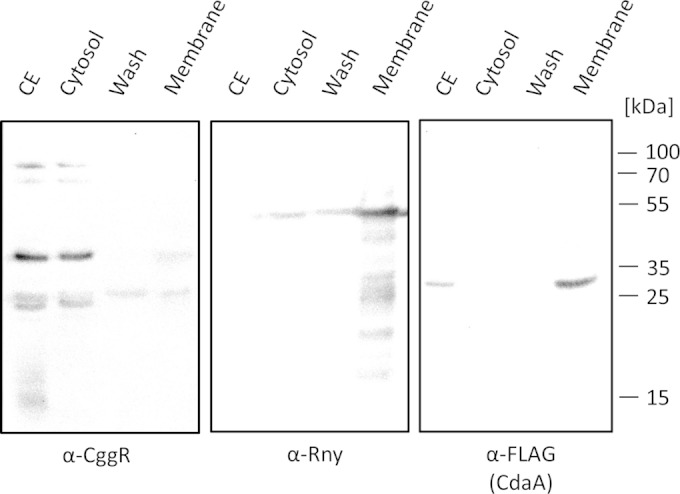
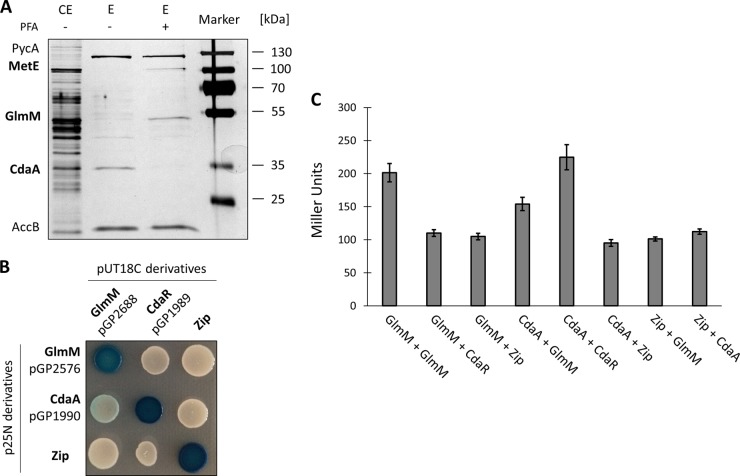
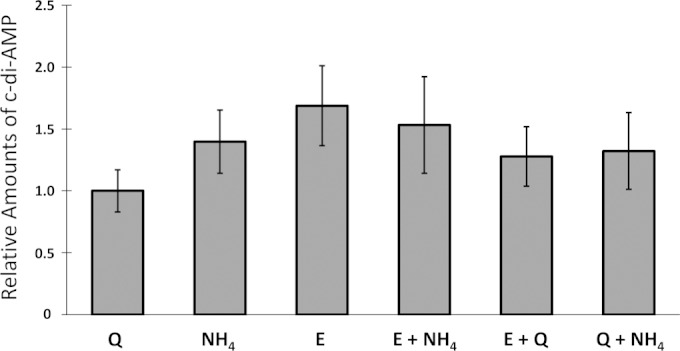
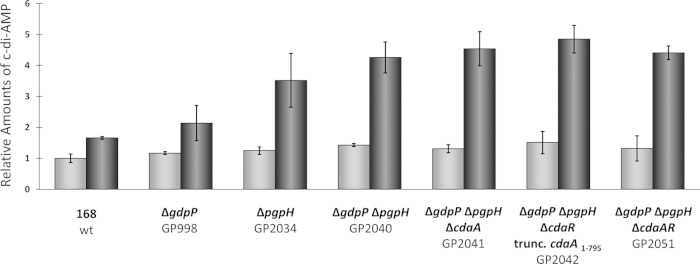
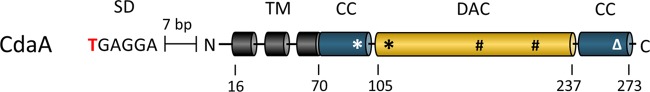
Gram-positive bacteria synthesize the second messenger cyclic di-AMP (c-di-AMP) to control cell wall and potassium homeostasis and to secure the integrity of their DNA. In the firmicutes, c-di-AMP is essential for growth. The model organism Bacillus subtilis encodes three diadenylate cyclases and two potential phosphodiesterases to produce and degrade c-di-AMP, respectively. Among the three cyclases, CdaA is conserved in nearly all firmicutes, and this enzyme seems to be responsible for the c-di-AMP that is required for cell wall homeostasis. Here, we demonstrate that CdaA localizes to the membrane and forms a complex with the regulatory protein CdaR and the glucosamine-6-phosphate mutase GlmM. Interestingly, cdaA, cdaR, and glmM form a gene cluster that is conserved throughout the firmicutes. This conserved arrangement and the observed interaction between the three proteins suggest a functional relationship. Our data suggest that GlmM and GlmS are involved in the control of c-di-AMP synthesis. These enzymes convert glutamine and fructose-6-phosphate to glutamate and glucosamine-1-phosphate. c-di-AMP synthesis is enhanced if the cells are grown in the presence of glutamate compared to that in glutamine-grown cells. Thus, the quality of the nitrogen source is an important signal for c-di-AMP production. In the analysis of c-di-AMP-degrading phosphodiesterases, we observed that both phosphodiesterases, GdpP and PgpH (previously known as YqfF), contribute to the degradation of the second messenger. Accumulation of c-di-AMP in a gdpP pgpH double mutant is toxic for the cells, and the cells respond to this accumulation by inactivation of the diadenylate cyclase CdaA.
Importance: Bacteria use second messengers for signal transduction. Cyclic di-AMP (c-di-AMP) is the only second messenger known so far that is essential for a large group of bacteria. We have studied the regulation of c-di-AMP synthesis and the role of the phosphodiesterases that degrade this second messenger. c-di-AMP synthesis strongly depends on the nitrogen source: glutamate-grown cells produce more c-di-AMP than glutamine-grown cells. The accumulation of c-di-AMP in a strain lacking both phosphodiesterases is toxic and results in inactivation of the diadenylate cyclase CdaA. Our results suggest that CdaA is the critical diadenylate cyclase that produces the c-di-AMP that is both essential and toxic upon accumulation.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Cyclic-di-AMP synthesis by the diadenylate cyclase CdaA is modulated by the peptidoglycan biosynthesis enzyme GlmM in Lactococcus lactis.Mol Microbiol. 2016 Mar;99(6):1015-27. doi: 10.1111/mmi.13281. Epub 2015 Dec 15. Mol Microbiol. 2016. PMID: 26585449
-
Cyclic di-AMP homeostasis in bacillus subtilis: both lack and high level accumulation of the nucleotide are detrimental for cell growth.J Biol Chem. 2013 Jan 18;288(3):2004-17. doi: 10.1074/jbc.M112.395491. Epub 2012 Nov 28. J Biol Chem. 2013. PMID: 23192352 Free PMC article.
-
Phenotypes Associated with the Essential Diadenylate Cyclase CdaA and Its Potential Regulator CdaR in the Human Pathogen Listeria monocytogenes.J Bacteriol. 2015 Nov 2;198(3):416-26. doi: 10.1128/JB.00845-15. Print 2016 Feb 1. J Bacteriol. 2015. PMID: 26527648 Free PMC article.
-
Replenishing the cyclic-di-AMP pool: regulation of diadenylate cyclase activity in bacteria.Curr Genet. 2016 Nov;62(4):731-738. doi: 10.1007/s00294-016-0600-8. Epub 2016 Apr 13. Curr Genet. 2016. PMID: 27074767 Review.
-
Making and Breaking of an Essential Poison: the Cyclases and Phosphodiesterases That Produce and Degrade the Essential Second Messenger Cyclic di-AMP in Bacteria.J Bacteriol. 2018 Dec 7;201(1):e00462-18. doi: 10.1128/JB.00462-18. Print 2019 Jan 1. J Bacteriol. 2018. PMID: 30224435 Free PMC article. Review.
Cited by
-
Control of asparagine homeostasis in Bacillus subtilis: identification of promiscuous amino acid importers and exporters.J Bacteriol. 2024 Feb 22;206(2):e0042023. doi: 10.1128/jb.00420-23. Epub 2024 Jan 9. J Bacteriol. 2024. PMID: 38193659 Free PMC article.
-
A decade of research on the second messenger c-di-AMP.FEMS Microbiol Rev. 2020 Nov 24;44(6):701-724. doi: 10.1093/femsre/fuaa019. FEMS Microbiol Rev. 2020. PMID: 32472931 Free PMC article. Review.
-
Sustained sensing in potassium homeostasis: Cyclic di-AMP controls potassium uptake by KimA at the levels of expression and activity.J Biol Chem. 2019 Jun 14;294(24):9605-9614. doi: 10.1074/jbc.RA119.008774. Epub 2019 May 6. J Biol Chem. 2019. PMID: 31061098 Free PMC article.
-
All DACs in a Row: Domain Architectures of Bacterial and Archaeal Diadenylate Cyclases.J Bacteriol. 2023 Apr 25;205(4):e0002323. doi: 10.1128/jb.00023-23. Epub 2023 Apr 6. J Bacteriol. 2023. PMID: 37022175 Free PMC article. Review.
-
Cyclic di-AMP, a multifaceted regulator of central metabolism and osmolyte homeostasis in Listeria monocytogenes.Microlife. 2023 Jan 28;4:uqad005. doi: 10.1093/femsml/uqad005. eCollection 2023. Microlife. 2023. PMID: 37223746 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
