

Accelerated evolution of the ASPM gene controlling brain size begins prior to human brain expansion
- PMID: 15045028
- PMCID: PMC374243
- DOI: 10.1371/journal.pbio.0020126
Accelerated evolution of the ASPM gene controlling brain size begins prior to human brain expansion
Abstract
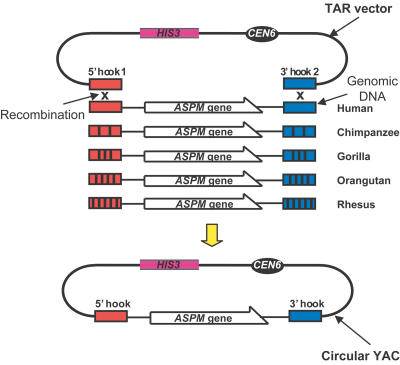
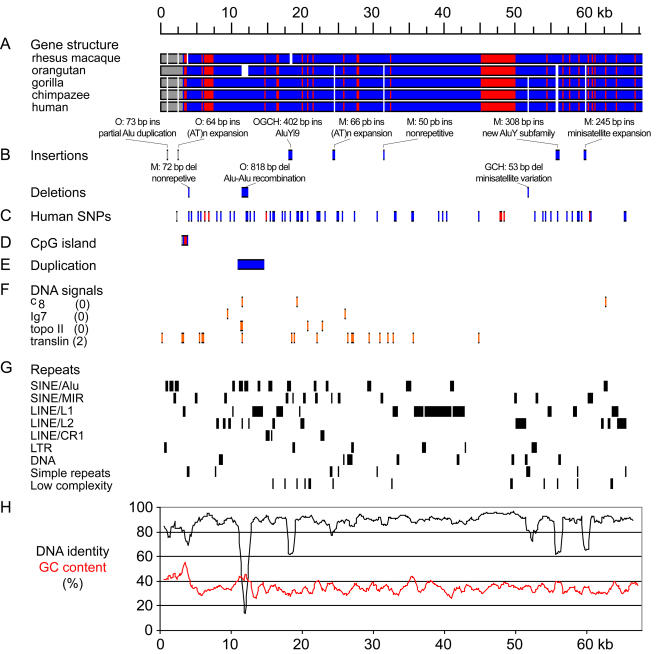
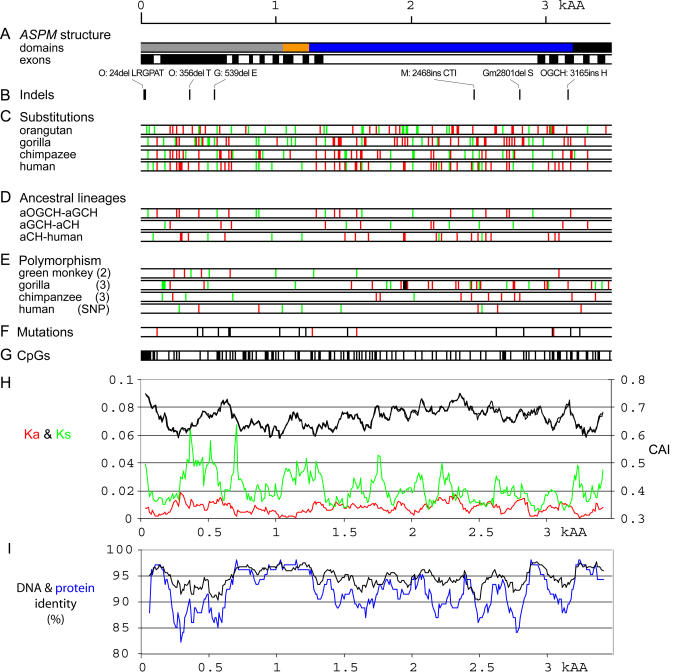
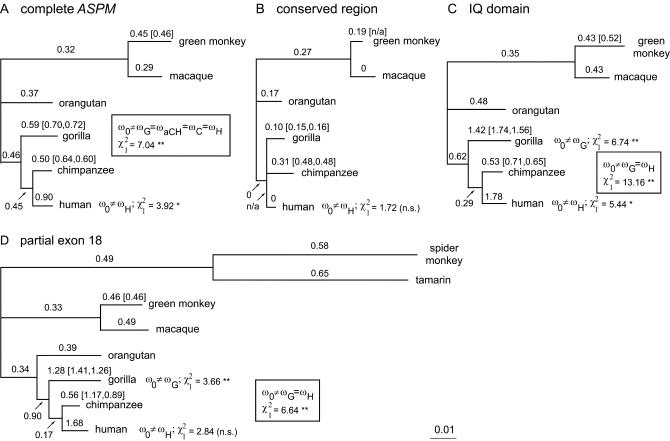
Primary microcephaly (MCPH) is a neurodevelopmental disorder characterized by global reduction in cerebral cortical volume. The microcephalic brain has a volume comparable to that of early hominids, raising the possibility that some MCPH genes may have been evolutionary targets in the expansion of the cerebral cortex in mammals and especially primates. Mutations in ASPM, which encodes the human homologue of a fly protein essential for spindle function, are the most common known cause of MCPH. Here we have isolated large genomic clones containing the complete ASPM gene, including promoter regions and introns, from chimpanzee, gorilla, orangutan, and rhesus macaque by transformation-associated recombination cloning in yeast. We have sequenced these clones and show that whereas much of the sequence of ASPM is substantially conserved among primates, specific segments are subject to high Ka/Ks ratios (nonsynonymous/synonymous DNA changes) consistent with strong positive selection for evolutionary change. The ASPM gene sequence shows accelerated evolution in the African hominoid clade, and this precedes hominid brain expansion by several million years. Gorilla and human lineages show particularly accelerated evolution in the IQ domain of ASPM. Moreover, ASPM regions under positive selection in primates are also the most highly diverged regions between primates and nonprimate mammals. We report the first direct application of TAR cloning technology to the study of human evolution. Our data suggest that evolutionary selection of specific segments of the ASPM sequence strongly relates to differences in cerebral cortical size.
Conflict of interest statement
The authors have declared that no conflicts of interest exist.
Figures
Similar articles
-
Adaptive evolution of ASPM, a major determinant of cerebral cortical size in humans.Hum Mol Genet. 2004 Mar 1;13(5):489-94. doi: 10.1093/hmg/ddh055. Epub 2004 Jan 13. Hum Mol Genet. 2004. PMID: 14722158
-
Genetic basis of brain size evolution in cetaceans: insights from adaptive evolution of seven primary microcephaly (MCPH) genes.BMC Evol Biol. 2017 Aug 29;17(1):206. doi: 10.1186/s12862-017-1051-7. BMC Evol Biol. 2017. PMID: 28851290 Free PMC article.
-
Aspm knockout ferret reveals an evolutionary mechanism governing cerebral cortical size.Nature. 2018 Apr;556(7701):370-375. doi: 10.1038/s41586-018-0035-0. Epub 2018 Apr 11. Nature. 2018. PMID: 29643508 Free PMC article.
-
Evolution of primary microcephaly genes and the enlargement of primate brains.Curr Opin Genet Dev. 2005 Jun;15(3):241-8. doi: 10.1016/j.gde.2005.04.009. Curr Opin Genet Dev. 2005. PMID: 15917198 Review.
-
Autosomal recessive primary microcephaly (MCPH): a review of clinical, molecular, and evolutionary findings.Am J Hum Genet. 2005 May;76(5):717-28. doi: 10.1086/429930. Epub 2005 Mar 31. Am J Hum Genet. 2005. PMID: 15806441 Free PMC article. Review.
Cited by
-
Aspm specifically maintains symmetric proliferative divisions of neuroepithelial cells.Proc Natl Acad Sci U S A. 2006 Jul 5;103(27):10438-10443. doi: 10.1073/pnas.0604066103. Epub 2006 Jun 23. Proc Natl Acad Sci U S A. 2006. PMID: 16798874 Free PMC article.
-
Analysis of oncogenic signaling networks in glioblastoma identifies ASPM as a molecular target.Proc Natl Acad Sci U S A. 2006 Nov 14;103(46):17402-7. doi: 10.1073/pnas.0608396103. Epub 2006 Nov 7. Proc Natl Acad Sci U S A. 2006. PMID: 17090670 Free PMC article.
-
Phylogenomic analyses reveal convergent patterns of adaptive evolution in elephant and human ancestries.Proc Natl Acad Sci U S A. 2009 Dec 8;106(49):20824-9. doi: 10.1073/pnas.0911239106. Epub 2009 Nov 19. Proc Natl Acad Sci U S A. 2009. PMID: 19926857 Free PMC article.
-
Detecting natural selection by empirical comparison to random regions of the genome.Hum Mol Genet. 2009 Dec 15;18(24):4853-67. doi: 10.1093/hmg/ddp457. Epub 2009 Sep 25. Hum Mol Genet. 2009. PMID: 19783549 Free PMC article.
-
A Likelihood Approach for Uncovering Selective Sweep Signatures from Haplotype Data.Mol Biol Evol. 2020 Oct 1;37(10):3023-3046. doi: 10.1093/molbev/msaa115. Mol Biol Evol. 2020. PMID: 32392293 Free PMC article.
References
-
- Abeysinghe SS, Chuzhanova N, Krawczak M, Ball EV, Cooper DN. Translocation and gross deletion breakpoints in human inherited disease and cancer. I: Nucleotide composition and recombination-associated motifs. Hum Mutat. 2003;22:229–244. - PubMed
-
- Bernardi G, Bernardi G. Codon usage and genome composition. J Mol Evol. 1985;22:363–365. - PubMed
-
- Bond J, Roberts E, Mochida GH, Hampshire DJ, Scott S, et al. ASPM is a major determinant of cerebral cortical size. Nat Genet. 2002;32:316–320. - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
